Invertebrate Anatomy OnLine
Vaejovis carolinianus ©
Carolina scorpion
29may2007
Copyright 2001 by
Richard Fox
Preface
This is one of many exercises available from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises can be accessed by clicking on the links to the left. A glossary and chapters on supplies and laboratory techniques are also. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Arthropoda P, Chelicerata sP, Euchelicerata, Arachnida C, Scorpiones O, Vaejovidae F (Fig 16-15, 18-47)
Arthropoda P
Arthropoda, by far the largest and most diverse animal taxon, includes chelicerates, insects, myriapods, and crustaceans as well as many extinct taxa such as Trilobitomorpha. The segmented body primitively bears a pair of jointed appendages on each segment. The epidermis secretes a complex cuticular exoskeleton which must be molted to permit increase in size. Extant arthropods exhibit regional specialization in the structure and function of segments and appendages but the ancestor probably had similar appendages on all segments. The body is typically divided into a head and trunk, of which the trunk is often further divided into thorax and abdomen.
The gut consists of foregut, midgut, and hindgut and extends the length of the body from anterior mouth to posterior anus. Foregut and hindgut are epidermal invaginations, being derived from the embryonic stomodeum and proctodeum respectively, and are lined by cuticle, as are all epidermal surfaces of arthropods. The midgut is endodermal and is responsible for most enzyme secretion, hydrolysis, and absorption.
The coelom is reduced to small spaces associated with the gonads and kidney. The functional body cavity is a spacious hemocoel divided by a horizontal diaphragm into a dorsal pericardial sinus and a much larger perivisceral sinus. Sometimes there is a small ventral perineural sinus surrounding the ventral nerve cord.
The hemal system includes a dorsal, contractile, tubular, ostiate heart that pumps blood to the hemocoel. Excretory organs vary with taxon and include Malpighian tubules, saccate nephridia, and nephrocytes. Respiratory organs also vary with taxon and include many types of gills, book lungs, and tracheae.
The nervous system consists of a dorsal, anterior brain of two or three pairs of ganglia, circumenteric connectives, and a paired ventral nerve cord with segmental ganglia and segmental peripheral nerves. Various degrees of condensation and cephalization are found in different taxa.
Development is derived with centrolecithal eggs and superficial cleavage. There is frequently a larva although development is direct in many. Juveniles pass through a series of instars separated by molts until reaching the adult size and reproductive condition. At this time molting and growth may cease or continue, depending on taxon.
Chelicerata sP
Chelicerata is a large taxon that includes spiders, scorpions, pseudoscorpions, ticks, mites, horseshoe crabs, sea spiders, and many others. The group originated in marine habitats but almost all modern chelicerates are terrestrial.
The body is divided into an anterior cephalothorax with six pairs of appendages and a posterior abdomen which, in most groups, does not bear appendages or has highly modified appendages. The first appendages of the cephalothorax are the chelicerae. Antennae are not present and the brain has two regions rather than the three found in mandibulates. Appendages are primitively biramous but are uniramous in almost all Recent taxa.
Euchelicerata
The segments of the cephalothorax are fused and covered by a dorsal shield, or carapace. Two median eyes are present.
Arachnida C
Arachnids are the terrestrial descendents of the early aquatic chelicerates. The taxon includes the mites, scorpions, pseudoscorpions, spiders, harvestmen, and several other taxa. All are adapted for a terrestrial existence with internal gas exchange and a waterproof integument. Some are capable of silk production and many use toxins to subdue or kill the prey. Most are carnivores, digestion is usually outside the body, and food is liquefied before ingestion.
Scorpiones O
Scorpions are primitive, ancient chelicerates that have changed little since the Silurian Period. The body consists of a segmented cephalothorax of six segments and an abdomen of twelve. The cephalothorax is covered by a carapace and bears chelicerae, pedipalps, and four pairs of walking legs. Scorpions are nocturnal predators feeding primarily on insects and other arthropods. Digestion begins externally and is completed intracellularly. The chelicerae and pedipalps are chelate and the distinctive pedipalps are large claws. The telson is a venomous sting. The second abdominal segment bears a pair of comb-like pectines.
The primitive nervous system is annelid-like with little cephalization. The heart is a dorsal tube in the anterior abdomen. Excretion is accomplished by saccate nephridia (coxal glands) and Malpighian tubules. Respiration is via four pairs of book lungs and the respiratory pigment hemocyanin is present in solution in the blood. There is a variety of sense organs. Poison from glands in the sting is injected into the prey.
Scorpions are gonochoric with complex mating behavior resulting in transfer of spermatophores from male to female. Fertilization is internal without copulation. Scorpions are viviparous and the female may care for the juveniles after birth.
Laboratory Specimens
Living scorpions are not available in large numbers, are difficult to handle safely, and are expensive when purchased. Some biological supply companies will provide them, however, and sometimes very large specimens can be purchased at pet stores. In either case, they are usually too expensive for use in most zoology courses, except perhaps as demonstrations. For these reasons it is assumed that preserved specimens will be used for this exercise but if living material is available, so much the better.
This exercise is written for the Carolina scorpion, Vaejovis carolinianus , which is one of the species provided commercially by biological supply companies in the North America. This is the only native scorpion in most of the eastern United States. It is a southeastern species reported from the Carolinas, Kentucky, Tennessee, Alabama, Mississippi, and Georgia and was originally described from South Carolina in 1821. Scorpion diversity is much higher in arid habitats in the western United States with 11 species in California and 8 in Arizona. Although diversity is highest in deserts, scorpions are also found in forests and grasslands.
Some common North American scorpions and their families are: Buthidae; Centruroides (including the lethal species C. sculptuatus, which killed 65 people in Arizona between 1929 and 1948), Vaejovidae; Vaejovis (the most common North American scorpions), Hadrurus (including H. hirstutus, the largest North American species), andAnuroctonus, Chactidae; Superstitiona. Most scorpions conform to a common body plan so it makes little difference which species is used. Specimens provided by biological supply houses are not identified.
Because of the small size of the scorpions available commercially, their study in a laboratory exercise is usually limited to external anatomy. Internal anatomy is covered by this exercise, however, for those who wish to pursue their study further. Conduct the study with a dissecting microscope with the specimen immersed in water or alcohol.
External Anatomy
Tagmata
Scorpion body segments are grouped into regions, or tagmata, consisting of the anterior cephalothorax, or prosoma, and the posterior abdomen, or opisthosoma (Figs 1, 2, 18-10, 18-11). The abdomen is further divisible into a wide anterior preabdomen (= mesosoma) and a narrow, posterior postabdomen (= metasoma). The segments of the cephalothorax and preabdomen bear paired segmental appendages, or their derivatives, but the postabdomen has no appendages or appendage derivatives.
The exoskeletal covering of a typical arthropod segment is divided into a dorsal tergite, a ventral sternite, and two lateral pleurites. These regions may be hardened sclerites or they may be soft and flexible articular membranes permitting motion between sclerites. This basic plan is present in the scorpions but with regional modification, especially anteriorly.
>1a. The scorpion cuticle fluoresces green under ultraviolet light. If a living specimen is available in the laboratory, view it illuminated with ultraviolet light in a darkened room. (Warning: UV radiation can damage your retina if the source is viewed directly.) Preserved specimens also fluoresce but not nearly so strongly as living animals. Expose an insect or spider to the UV light for comparison. <
Figure 1. Dorsal view of the scorpion, Vaejovis carolinianus. Scorpion40La.gif
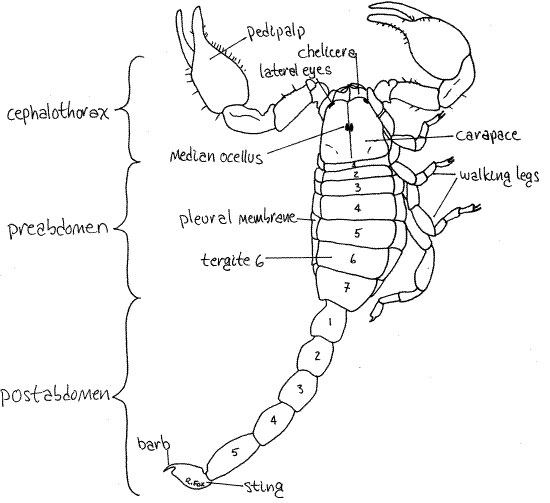
Cephalothorax
Examine the dorsal surface. The adult cephalothorax consists of six segments covered dorsally by a single, unsegmented, sclerotized carapace (Fig 1, 18-10). The underlying segmentation of the cephalothorax is not apparent dorsally but is obvious ventrally. There is a pair of median ocelli on the dorsal midline of the carapace and their lenses are visible with higher magnification (25X). In Vaejovis several pairs of small lateral eyes are present on the antero-lateral corners of the carapace.
The first pair of cephalothoracic appendages is the chelate chelicerae (Figs 1, 2). These are short, stout pincers extending anteriorly from under the anterior margin of the carapace. Their distal two articles form a chela, or pincer, used to hold the prey and tear prey into small pieces. The chelicerae form the roof of the preoral cavity, a space enclosed by the bases of the anterior appendages in which digestion of food begins prior to ingestion (Fig 18-12).
Figure 2. Ventral view of the scorpion, Vaejovis carolinianus. Scorpion41L.gif
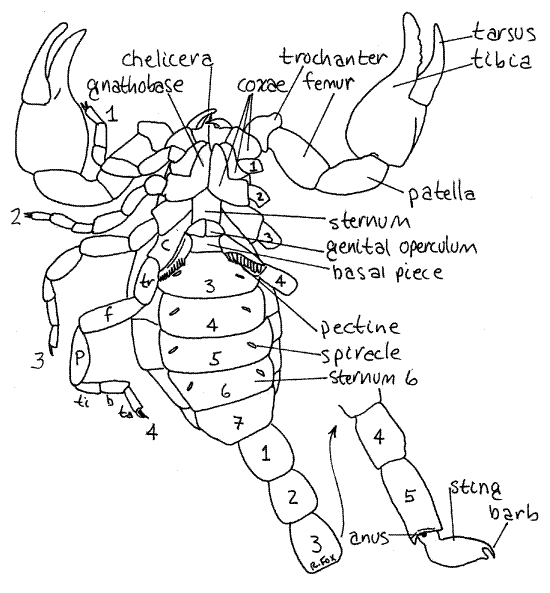
The second pair of appendages is the large, prehensile, pedipalps so characteristic of scorpions (Fig 2). Like the chelicerae, the pedipalps are chelate with the last two articles working together in a prehensile pinching mechanism. The pedipalps are used in prey capture and defense and bear fine sensory setae known as trichobothria.
Each pedipalp is composed of six articles. In order, from proximal to distal, they are the coxa, trochanter, femur, patella, tibia, and tarsus (Fig 2). The tibia is the large, swollen penultimate article and the tarsus is the slender movable finger of the chela. The immovable finger of the chela is a process on the tibia. The coxae of the pedipalps form the sides of the preoral cavity.
The next four segments each bear a pair of walking legs. Each walking leg comprises eight articles which are, in order from proximal to distal, coxa, trochanter, femur, patella, tibia, basitarsus, tarsus, and apotele. The apotele is often so small as to be indiscernible. The tarsus bears a pair of distal tarsal claws. Note the flexible, unsclerotized articular membranes between the articles of the limbs.
Look at the legs in ventral view. The coxae of the walking legs contribute importantly to the ventral surface of the cephalothorax. The coxae of the first two walking legs completely replace the ancestral sternites as the hardened plates of the venter of the anterior cephalothorax. The right and left coxae touch each other on the ventral midline with no sternites between them. These coxae have platelike anterior expansions known as gnathobases which extend anteriorly to form the floor of the preoral cavity (Fig 2).
Food is captured by the pedipalps, torn apart and held in the preoral cavity by the chelicerae, and masticated by the gnathobases. Hydrolytic enzymes flood onto the food from the midgut, hydrolysis begins before ingestion, and the resulting liquids are sucked into the gut. A setal filter on the gnathobases excludes particulates from entry into the gut.
The coxae of walking legs 3 and 4 are not expanded in this manner and do not meet on the midline. The sternites of their two segments are present but fused together to form a triangular or pentagonal plate known simply as the sternum. It lies on the ventral midline of the posterior cephalothorax. The coxae of legs 3 and 4 attach to its lateral borders.
Abdomen
The remainder of the body is the abdomen. The seven wide anterior abdominal segments are the preabdomen (Figs 1, 2, 18-11). The narrow posterior five segments are the tail-like postabdomen. The sting (telson) is at the posterior end of the postabdomen.
Ventrally, the first two preabdominal segments have modified sternites. On the ventral midline of the first segment is a pair of genital opercula (Fig 2) which cover the gonopore. The two opercula of females are separate whereas those of males are fused to form a single plate. What sex is your specimen? _________ The opercula are derived from the appendages of this segment.
The second preabdominal segment has a sternite known as the basal piece and a pair of unique, comb-like, sensory pectines derived from the segmental appendages of this segment. The pectines are sensory structures with chemo- and mechanoreceptors. Their role is unclear but they may be used to recognize appropriate sites for spermatophore deposition.
Each of the five remaining preabdominal segments has a large unmodified sternite. Dorsally each preabdominal segment is covered by a sclerotized tergite (Fig 1). Laterally the sternites and tergites of each segment are united by a broad, unsclerotized, flexible pleural membrane (Fig 2).
Preabdominal segments 3-6 each bear a pair of ventral spiracles, each of which opens into a book lung (Fig 2, 18-11). The spiracles are usually oblique slits on the ventrolateral surface of the sternite but they may be other shapes in some species and they may be difficult to see in preserved material. If so, the best way to find them is to probe the likely area gently with a microneedle. The book lungs, which may be faintly visible through the exoskeleton, are derivatives of four pairs of segmental appendages. The seventh preabdominal segment has no appendage derivatives.
The postabdomen consists of five short, cylindrical segments whose tergites and sternites are fused to form an unbroken cuticular ring. There are no appendages or their derivatives on the postabdomen and no pleural membranes. The posterior unit of the postabdomen is the sting (telson), which is not a true segment. It bears the hollow, terminal barb at its tip. Two poison glands are housed in the swollen portion of the sting. The neurotoxin produced by the glands empties via ducts to a pore at the tip of the barb. The sting is usually used in defense or to quell struggling prey and that of most scorpions is unpleasant but not life-threatening to humans but a few, mostly in the family Buthidae, are lethal.
The ventral anus opens in the articulating membrane between segment five and the base of the sting. It is on the ventral midline at the posterior end of segment five.
Internal Anatomy
Most scorpions available in teaching laboratories are too small for detailed study of internal anatomy. The difficulty is compounded in preserved specimens as they are very brittle. A superficial study is possible, however, and may be undertaken if desired. The dissection should be conducted under water using a dissecting microscope.
" Place the scorpion dorsal side up in a small dissecting pan of water. An anchovy can makes an ideal size (see Techniques chapter). Use # 1 insect pins through the femora of the pedipalps and last walking legs to anchor the animal firmly to the wax.
Use fine scissors to make a lateral, longitudinal incision through the pleural membranes on both sides of the preabdomen. Begin with the seventh (last) preabdominal segment and cut anteriorly for the length of the preabdomen. Make transverse cuts across the body in the articulating membranes between the preabdomen and postabdomen and between the cephalothorax and preabdomen. Carefully lift the dorsal body wall of the preabdomen and cut the muscles holding it in place so you can remove it. There are seven pairs of dorso-ventral muscles attached to the tergites of the preabdomen and these must be cut in order to remove the body wall. Be very careful that you do not damage the heart which lies on the dorsal midline immediately inside the dorsal body wall. Look at its inside surface of the roof of the preabdomen and note the longitudinal muscles running from tergite to tergite.
Now use the fine scissors to cut the pleural membrane on both sides between the carapace and the bases of the cephalothoracic appendages. Lift the carapace a little and cut the muscles attached to its inner surface. The cephalothorax contains numerous large, powerful, extrinsic muscles that operate the appendages. Remove the carapace. The median eyes will probably remain attached to the body. Note the muscles in the interior of the cephalothorax. The largest are the extrinsic muscles of the pedipalps and chelicerae.
Preserved scorpions do not lend themselves to systematic, orderly study by organ systems. Instead, consider the structures and organs in the order in which they are encountered.
Hemal System
The space you have revealed in the interior of the cephalothorax and preabdomen is the hemocoel (Fig 3). It is a space in the connective tissue compartment and is part of the hemal system. It is the functional body cavity and the visceral organs are contained within it, where they are bathed in blood, or hemolymph. The hemocoel is partitioned by a vertical, transverse, membranous diaphragm between the cephalothorax and preabdomen (Fig 3).
The dorsal, tubular heart extends the length of the preabdomen (Fig 3) and bears seven pairs of ostia. These are pores in the heart wall through which blood enters the heart lumen. Blood leaves the heart in anterior and posterior aortae and paired lateral arteries. Numerous paired alary muscles extend laterally from the walls of the heart and are responsible for expanding the heart lumen. Contraction is accomplished by circular muscles in the heart wall. The six pairs of dorsoventral muscles which you cut to remove the body wall lie beside the heart.
Figure 3. Dorsal view of the hemocoel of the cephalothorax and mesosome of Vaejovis. Scorpion42La.gif
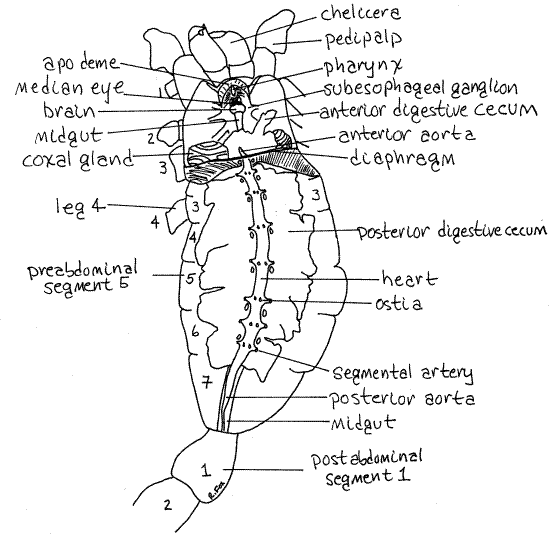
Anteriorly the heart turns ventrally, penetrates the diaphragm, and becomes the anterior aorta (Fig 3). The aorta then divides into many arteries to supply the cephalothorax with blood. Posteriorly the heart narrows to become the posterior aorta which enters the postabdomen (Fig 3).
Posterior Digestive System
The gut runs the length of the body from a mouth at the anterior end of the cephalothorax to the anus near the posterior end of the postabdomen (Fig 3). It is divided into an anterior foregut, middle midgut, and posterior hindgut. Little, if any, of it is visible at present. The midgut is a long tube covering most of the length of the cephalothorax, all of the preabdomen, and most of the postabdomen. The foregut and hindgut, at opposite ends of the midgut, are short and cuticularized. The midgut is a simple endodermal tube from which arise six pairs of diverticula, the digestive ceca. Intracellular digestion occurs in the digestive ceca.
These diverticula consist of five pairs of posterior digestive ceca in the preabdomen and one pair of anterior digestive ceca in the cephalothorax (Fig 3). The posterior ceca occupy most of the space in the preabdomen and are conspicuous. In preserved specimens they are clumped together and cannot usually be distinguished as separate entities. The anterior digestive ceca are in the posterior cephalothorax next to the diaphragm. Find the midgut where it emerges from the posterior end of the mass of digestive ceca.
Excretory System
Scorpions have a single pair of coxal glands in the fifth cephalothoracic segment (that of walking leg 3). Carefully move aside or remove the anterior digestive ceca on one side to reveal the pale coxal gland (= saccate nephridia) ventral to it (Fig 3). Be careful that the coxal gland does not come away with the digestive ceca. The swirling lines on its surface are suggestive of the gyri on the surface of the mammalian cerebrum. The coxal glands open through pores on the coxae of walking leg 3. They concentrate and eliminate uric acid from the blood.
Two pairs of slender Malpighian tubules arise from the midgut at the posterior end of the preabdomen. They are usually overlooked in preserved specimens. One pair extends anteriorly through the diaphragm into the hemocoel of the cephalothorax. The other pair is in the hemocoel of the preabdomen. The Malpighian tubules concentrate and eliminate guanine.
Reproductive System
" Remove the heart and anterior aorta in the vicinity of the diaphragm. This will reveal the midgut where it penetrates the diaphragm ventral to the aorta. The midgut runs through the mass of posterior digestive ceca and then exits the preabdomen posteriorly to enter the postabdomen.
Carefully pick away pieces of the posterior digestive ceca from the preabdomen. This will reveal, and probably destroy in the process, the gonad and reproductive system.
In scorpions the gonad, whether ovary or testis, is a network large, longitudinal tubes embedded in the posterior digestive ceca of the preabdomen. The longitudinal tubes are connected by transverse tubes and with various vesicles arising from the tubes. The arrangement varies with species. The system opens at the gonopore on preabdominal segment 1 under the genital operculum. A more careful study of the reproductive system is not practical. Scorpions are viviparous. Fertilization is internal but without copulation. Instead, the female receives, by various means, a spermatophore from the male. Females retain fertilized eggs while they develop.
Respiratory System
The four pairs of book lungs are visible from the exterior as white areas situated laterally on the sternites of preabdominal segments 3-6 (Fig 2, 18-11). The lungs contain large numbers of cuticular folds, or lamellae, resembling the pages of a book, which provide surface area for respiration (Fig 18-6). Gasses are exchanged between air and blood across the thin cuticle of the lamellae.
" Use fine scissors to make a cross section across one of the book lungs, cutting through the body wall to do so. Spread the inner and outer surfaces of the lung apart with microneedle and forceps and look at the cut edge with 25X or higher. This will reveal the thin cuticular lamellae of the lung.
Anterior Digestive System
Follow the gut anteriorly from the diaphragm, carefully removing connective tissue to do so. It passes through the large circumesophageal nerve ring (Fig 3, 4). The nerve ring lies ventral to the median ocellus and will be discussed shortly. Do not damage it.
Figure 4. The interior of the cephalothorax of Vaejovis. Scorpion43L.gif
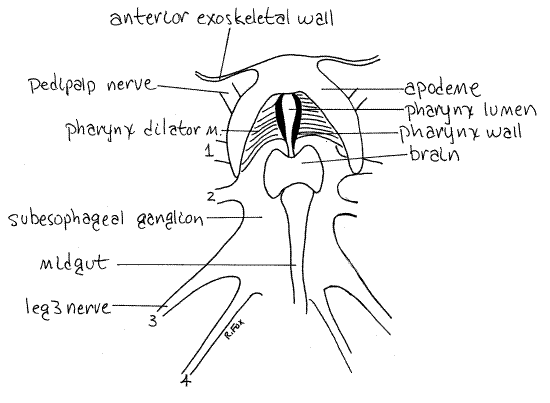
From the nerve ring anteriorly the gut is foregut and consists of the pharynx and esophagus. The short esophagus passes through the nerve ring and obscured by it (Figs 3, 4). The pharynx is anterior to the nerve ring. Look on the inside surface of the anterior wall of the cephalothorax, between the two chelicerae, for a large U-shaped apodeme (Figs 3, 4). The pharynx lies on the midline between the two arms of the "U" of the apodeme.
A pair of short, stout pharynx dilator muscles originate on the insides of the arms of the apodeme and insert on the elastic, cuticular walls of the pharynx (Fig 4). These walls are curved and look like a pair of closely spaced parentheses (). Juices from the masticated, pre-digested food in the preoral cavity are sucked through the mouth into the pharynx by contractions of the dilator muscles.
The mouth is immediately anterior to the pharynx but you will not see it. From the pharynx you can slip a microneedle into it however. The short esophagus passes posteriorly through the nerve ring and connects the pharynx with the midgut. The midgut is a simple, long, narrow tube with digestive ceca that extends most of the length of the animal. The posterior end of the gut is the hindgut, or rectum, in the posterior postabdomen.
Nervous System
Relocate the circumesophageal nerve ring. The brain is the part of the nerve ring dorsal to the esophagus immediately posterior to the pharynx (Fig 4). The median ocellus is dorsal to it. The chelicerate brain consists of protocerebrum and tritocerebrum but you cannot distinguish them in gross dissection. The protocerebrum innervates the eyes and the tritocerebrum, the chelicerae. You probably will not see these nerves. Two short stout circumesophageal connectives exit the lateral margins of the brain and pass around the esophagus to join the subesophageal ganglion ventral to the gut (Figs 3, 4, 18-7). The connectives adhere tightly to the esophagus. The large subesophageal ganglion is wide and flat, covering much of the floor of the center of the cephalothorax. It comprises the ganglia of the pedipalps, four pairs of walking legs, and the first four preabdominal segments (Fig 18-7). If you carefully remove the connective tissue and muscles dorsal to the ganglion, you can see the larger nerves that exit it. Those to the pedipalps and walking legs are largest and are usually visible.
A pair of longitudinal nerve cords exits the posterior end of the ganglionic mass and passes posteriorly as the ventral nerve cord lying on the floor of the preabdomen and postabdomen. It has three ganglia in the preabdomen and four in the postabdomen (Fig 18-7). The nerve cord and its ganglia are difficult to demonstrate in preserved specimens.
References
Awati PR, Tembe VB . 1956. Buthus tamulus (Fabr.) The Indian scorpion; morphology, anatomy, and bionomics. Univ. Bombay, Zool. Mem. 2:1-62.
Benton CLB . 1973. Studies on the biology and ecology of the scorpion, Vejovis carolinianus (Beauvois). Ph.D diss., Dept. Biology, Univ. Alabama. (Univ. Microfilms, 320 N. Zeeb Rd., Ann Arbor, MI, 48106.)
Cheng-Pin P. 1940. Morphology and anatomy of the Chinese scorpion, Buthus martensi Karsh. Peking Nat. Hist. Bull. 14:103-117, pls. 1-2.
Kaestner A. 1967. Invertebrate Zoology, vol. II. Arthropod Relatives, Chelicerata, Myriapoda. Wiley-Interscience, New York. 472p.
Polis GA. 1990. The Biology of Scorpions. Stanford Univ. Press, Palo Alto. 614p.
Savory TH. 1977. Arachnida 2 nd ed. Academic Press, New York. 340p.
Snodgrass RE . 1952. A Textbook of Arthropod Anatomy. Cornell, Ithaca. 363p.
Ruppert EE, Fox RS, Barnes RB. 2004. Invertebrate Zoology, A functional evolutionary approach, 7 th ed. Brooks Cole Thomson, Belmont CA. 963 pp.
Supplies
Dissecting microscope
Small dissecting pan (anchovy or sardine tin)
Preserved scorpion
#1 stainless steel dissecting pins