Invertebrate Anatomy OnLine
Popillia japonica ©
Japanese Beetle
30jun2006
Copyright 2001 by
Richard Fox
Lander University
Preface
This is one of many exercises available from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises can be accessed by clicking on the links in the column on the left. A glossary and chapters on supplies and laboratory techniques are also available. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Arthropoda P, Mandibulata sP, Tracheata, Hexapoda SC, Insecta C, Dicondylia, Pterygota, Metapterygota, Neoptera, Eumetabola, Holometabola, Coleoptera O, Polyphaga sO, Scarabaeoidea SF, Scarabaeidae F, Rutelinae sF, Anomalini T (Fig 16-15, 20-14, 20-15, 21-23)
Arthropoda P
Arthropoda, by far the largest and most diverse animal taxon, includes chelicerates, insects, myriapods, and crustaceans as well as many extinct taxa. The body is segmented and primitively bears a pair of jointed appendages on each segment. The epidermis secretes a complex cuticular exoskeleton which must be molted to permit increase in size. Extant arthropods exhibit regional specialization in the structure and function of segments and appendages. The body is typically divided into a head and trunk, of which the trunk is often itself divided into thorax and abdomen.
The gut consists of foregut, midgut, and hindgut and extends the length of the body from anterior mouth to posterior anus. Foregut and hindgut are epidermal invaginations, being derived from the embryonic stomodeum and proctodeum respectively, and are lined by cuticle, as are all epidermal surfaces. The midgut is endodermal and is responsible for most enzyme secretion, hydrolysis, and absorption.
The coelom is reduced to small spaces associated with the gonads and kidney. The functional body cavity is a spacious hemocoel divided by a horizontal diaphragm into a dorsal pericardial sinus and a much larger perivisceral sinus. Sometimes there is a small ventral perineural sinus surrounding the ventral nerve cord.
The hemal system includes a dorsal, contractile, tubular, ostiate heart that pumps blood to and from the hemocoel. Excretory organs vary with taxon and include Malpighian tubules, saccate nephridia, and nephrocytes. Respiratory organs also vary with taxon and include many types of gills, book lungs, and tracheae.
The nervous system consists of a dorsal, anterior brain of two or three pairs of ganglia, circumenteric connectives, and a paired ventral nerve cord with segmental ganglia and segmental peripheral nerves. Various degrees of condensation and cephalization are found in different taxa.
Development is derived with centrolecithal eggs and superficial cleavage. There is frequently a larva although development is direct in many. Juveniles pass through a series of instars separated by molts until reaching the adult size and reproductive condition. At this time molting and growth may cease or continue, depending on taxon.
Mandibulata sP
Mandibulata includes arthropods in which the third head segment bears a pair of mandibles. As currently conceived this taxon includes myriapods, hexapods, and crustaceans. Appendages may be uni- or biramous and habitats include marine, freshwater, terrestrial, and aerial.
Tracheata
Myriapods and hexapods share tracheae and a single pair of antennae and are sister taxa in Tracheata. Crustaceans, which have gills and lack tracheae, are excluded and form the sister group.
Hexapoda SC
The body is divided into three tagmata; head, thorax, and abdomen. Appendages are uniramous and a single pair of antennae is present. Three pairs of legs and two pairs of wings are found on the thorax of most adults. Hexapod legs are uniramous although there is increasing evidence that they evolved from multiramous appendages of their ancestors. Gas exchange is accomplished by trachea. Excretory organs are Malpighian tubules and the end product of nitrogen metabolism is uric acid. There is relatively little cephalization of the nervous system. Insects are gonochoric with copulation and internal fertilization.
Insecta C
Most hexapods are insects. A few hexapod taxa (orders) lack wings and have primitive mouthparts recessed into the head and belong to Entognatha, the sister taxon of Insecta. Insects have ectognath mouthparts and the adults (imagoes) of most taxa have wings.
Pterygota
The winged insects. These insects are derived from a winged common ancestor. Adults of most taxa have wings although they have been lost in some.
Eumetabola
Juveniles have no ocelli and there are six or fewer Malpighian tubules.
Holometabola
The final larval instar pupates and undergoes a radical metamorphosis in which it is converted to an imago, or adult. The imago is sexually mature and in most taxa has wings whereas larvae are immature and wingless. During metamorphosis many or most larval tissues are dismantled and adult structures built anew. Wings, for example, are manufactured from clusters of undifferentiated cells known as imaginal discs but not from preformed wingpads as in pauro- and hemimetabolous insects.
Coleoptera O
Coleoptera is the largest taxon (of equivalent rank) of organisms with about 300,000 described Recent species. The body of most beetles is heavily sclerotized. The forewings are sclerotized elytra, or wing covers, that do not participate in flight.
Natural History
The Japanese beetle, Popillia japonica, is an introduced species that may be all too abundant in summer in the eastern United States. It belongs to the scarab beetle family, Scaraebidae, and the shining leaf chafer subfamily. It was introduced into the United States from Japan in 1916. It is readily available alive (in season) in many locales and few will protest the loss of specimens for laboratory use. These are mandibulate insects and the herbivorous adults and larvae alike are serious pests in lawns, orchards, and gardens. The larvae, which are white grubs, live in the soil where they feed on the roots of overlying vegetation, especially turfgrass. Adults feed on the foliage and fruit of a variety of plants. They are univoltine (one generation/year) and diapause as grubs in the soil.
Laboratory Specimens
The exercise is an account of the external anatomy of imagoes but does not cover the internal anatomy. Adults are easily collected in the eastern United States in the summer by hand picking them from their host plants. They can be preserved in 40% isopropanol or 80% ethanol for later use during the academic year if so needed. The exercise can be conducted on anesthetized living specimens or alcoholic specimens. Anesthetization can be accomplished with carbon dioxide or exposure to a cotton ball dampened slightly with chloroform. Specimens can be killed by using ethyl acetate instead of chloroform on the cotton. Conduct the study with a dissecting microscope. The study is limited to external anatomy of adults. Another exercise in this series id devoted to the external anatomy of scarabaeid grubs. It is available under Cyclocephala in the column to the left.
External Anatomy
Place an anesthetized or preserved beetle in a small dissecting pan of tap water and examine it with the dissecting microscope at about 7X magnification. In spite of their evil reputation, these are really quite beautiful insects, especially when viewed with magnification. Note the green iridescence of the cuticle and the abundant white setae, especially on the ventral surface and in tufts on the sides of the abdomen.
Tagmata
The arthropod body is composed of a linear series of segments, some or all of which bear a pair of jointed segmental appendages. Most arthropods, including insects, are heteronomous, with segments and appendages variously modified and grouped in regions, or tagmata, specialized for different functions. The homonomous condition, with all segments and appendages alike over the length of the body, is primitive but, among Recent taxa, is approached only by a few crustaceans and no insects or chelicerates.
The three tagmata of the insect body are the anterior head, middle thorax, and posterior abdomen (Fig 21-1C). The head, which, other than its appendages, shows no external signs of segmentation, bears the eyes, antennae, mouth, and mouthparts (Fig 1, 2, 3). Its chief functions are sensory reception, neural integration, and feeding.
The thorax, whose primary function is locomotion, is larger and bears three pairs of legs and two pairs of wings.
The abdomen is usually the largest tagma and is conspicuously segmented. It houses most of the digestive, excretory, and reproductive viscera and appendages, when present, are specialized for copulation or oviposition. Most abdominal segments lack appendages and those present are highly modified. The external genitalia at the posterior end of the abdomen. are modified segmental appendages.
Figure 1. Dorsal view of the Japanese beetle, Popillia japonica. Beetle65L.gif
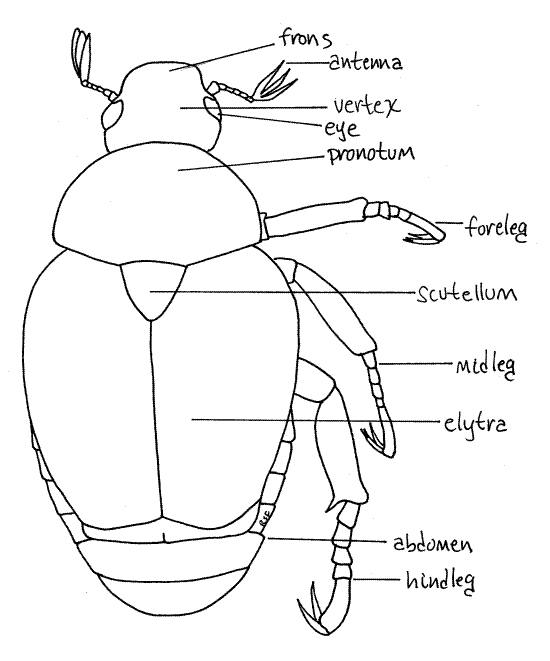
In beetles tagmatization is indistinct dorsally and the usual three insect tagmata are not readily apparent. Dorsally the head is visible but most of the thorax and abdomen are obscured by the large wing covers, or elytra which give a false impression of tagmatization. It is much easier to discern the three tagmata from the venter.
Head
The head is easy to distinguish, dorsally or ventrally (Fig 1, 2, 3, 21-1C, 21-14). It is enclosed in a heavily sclerotized head capsule (= cranium) that shows no conspicuous external signs of segmentation. It is the smallest of the three tagma and is well separated from the remainder of the body by the soft, flexible cervix, or neck (Fig 3). Ventrally the head fits neatly into a notch in the first segment of the thorax. Move the head back and forth to demonstrate the flexibility of the cervix. Find the conspicuous compound eyes on the sides of the head.
Figure 2. Ventral view of the head of Popillia japonica. Beetle66L.gif
The head capsule, or epicranium, is divided into regions, each of which represents a sclerite which has become fused with its neighbors to form the capsule (Fig 2). Sometimes, visible sutures remain between adjacent sclerites but these are poorly developed in Popillia. The top of the head, from the level of the eyes posteriorly, is the vertex. A fingerlike process of the vertex extends ventrally into the compound eye. The side of the head posterior to the eye is the gena. The posterior wall of the head capsule is the occiput. The occipital foramen (= foramen magnum) is a large hole in the center of the occiput. Through it pass the organ systems, such as nerve cord, gut, and musculature, connecting the head with the body. The foramen is covered by soft tissue but you can see its circular outline. Anterior to the vertex is the frons. In Popillia, the frons and vertex are separated by a faint suture. The frons (of Popillia) is shaped like a wide scoop with a ridge around the anterior margin. The head capsule turns ventrally anterior to the frons and forms a nearly vertical wall known as the clypeus (CLIP ee us) (Fig 3). The suture between the clypeus and the frons is the ridge mentioned previously. The mouthparts on the ventral surface of the head surround the mouth. These will be studied later.
Thorax
The thorax consists of three segments; the prothorax, mesothorax, and metathorax, but only the prothorax is visible in dorsal view. It is covered dorsally by a large oval, green sclerite, the pronotum (Fig 1, 21-14). Dorsally the remainder of the thorax and abdomen are hidden by the two large, rusty red wing covers, or elytra, arising from the second thoracic segment.
The three segments of the thorax are visible ventrally and each bears a pair of legs (Fig 3). The first segment is the prothorax with its prothoracic legs, or forelegs. The ventral sclerite of the prothorax is the prosternite. It is largely hidden by the large coxa of the foreleg.
The prothorax is connected with the mesothorax by a flexible articulation. Demonstrate this flexibility to yourself. Find the mesothoracic legs, or middle legs. The mesosternite is narrow and obscured by the long, narrow femur and tibia of the midlegs.
The third segment, the metathorax is rigidly articulated with the posterior border of the mesothorax. Demonstrate this fact. Together the fused meso- and metathorax are the pterothorax, so named because they bear the two pairs of wings (pter = wing) Identify the metathoracic legs, or hindlegs. The large conspicuous metasternite is easily found although it may appear to be part of the mesothorax rather than the metathorax. It bears a large, blunt metasternal process that points anteriorly.
Legs
Each of the three thoracic segments bears a pair of legs. The legs are easier to study later, after you have completed your study of the head.
Wings
Like most adult insects, beetles have two pairs of wings, one mesothoracic (forewings) and one metathoracic (hindwings). No insect has wings on the prothorax. Immature insects do not have wings and adults of some taxa also lack wings. Beetle forewings are heavy sclerotized and serve, not for flight, but as wing covers, or elytra, to protect the delicate hindwings, which do function in flight. At rest the elytra cover the membranous hindwings which are folded beneath them, out of harms way. Note the shield shaped scutellum, a conspicuous triangular sclerite between the anterior ends of the two elytra. Elytra are a Coleoptera apomorphy.
Use your finest forceps (with help from another forceps) to move one of the elytra aside so you can see the hindwings. The elytra move laterally and up. They may resist moving at first but after having been opened once they will move easily thereafter.
The hindwings, or metathoracic wings, are much longer than the elytra and consequently must be folded to fit in the space below the elytra. Carefully unfold one of the hindwings. Note its size, in relation to that of the elytra, and its delicacy. Find some of the sclerotized costae, or veins, that strengthen and support the wing.
Abdomen
The abdomen, consisting of five visible segments, is easily seen in ventral view. (In dorsal view, with the elytra and wings moved aside, you can see seven segments). The sternites of these segments are easy to find. Note the fringe of white setae on each sternite. From the dorsal aspect you can see the dorsal tergites of the most posterior abdominal segments. Move one of the elytra up and to the side to improve your view of the tergites. Each is a sclerified strip of dark cuticle extending transversely across the top of the abdomen. The fifth (posteriormost) tergite is triangular and is sometimes referred to as the pygidium.
Figure 3. Ventral view of Popillia. The head is pulled anteriorly to reveal the cervix. Similarly, the metasternite is elevated to reveal the anterior abdomen which is normally hidden. Beetle67L.gif
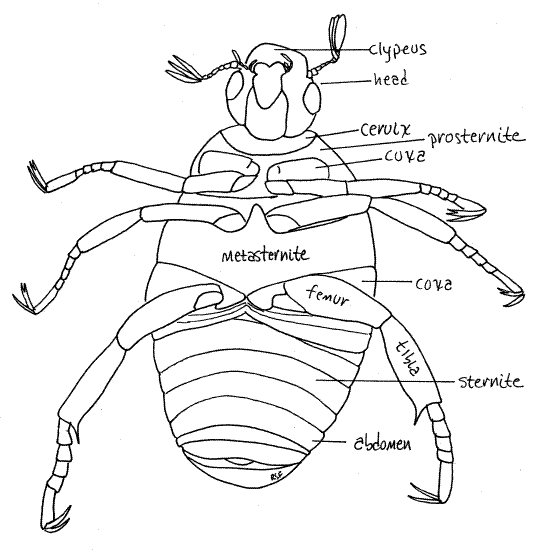
With the elytra still lifted and out of the way, find the lateral pleurites, which connect the tergites and sternites. These are soft, flexible, and pale creamy white. They are located on the lateral margins of the dorsum. Each pleurite has a small brownish spiracle.
While you are looking at the dorsal surface of the abdomen, count its segments. You will find that there are seven visible segments in dorsal view. The first two are hidden by the metasternite in ventral view so you could not see them.
Appendages
Head
Return now to the head and complete your study of it (Fig 2, 4). Increase the magnification to about 20X and look closely at the head. Grasp the prothorax with forceps and pull until it detaches from the mesothorax. Leave the head attached to the prothorax. This will make it easier to study the head.
Figure 4. En face view of Popillia japonica. Beetle68L.gif
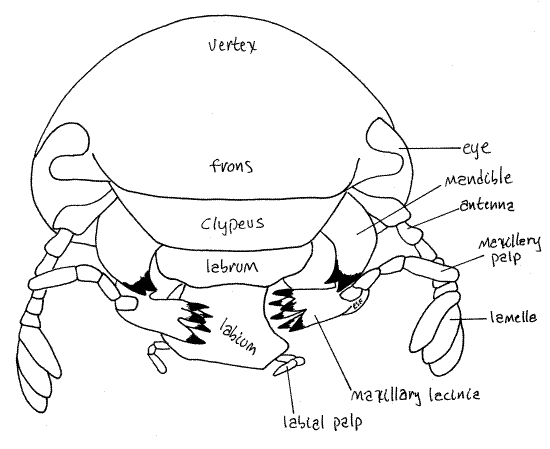
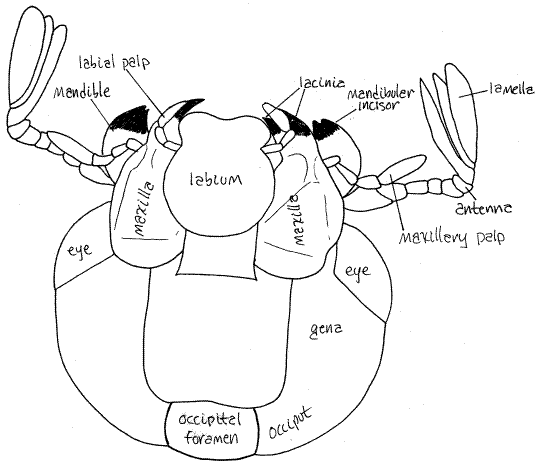
Antennae
The antennae arise on the anterior margin of the eye and are usually reflexed under the head in preserved specimens. Each antenna is composed of a linear series of six small articles (Fig 2, 4). Distally the last article bears three long, narrow, parallel plates, or lamellae. This type of antenna is said to be lamellate and it is characteristic of scarab beetles.
Labium
Tracheate arthropods, with the exception of millipedes and pauropods, have three pairs of mouthparts. The mouth is on the ventral surface of the head and opens from an external chamber, the preoral cavity, surrounded by the mouthparts. Study the mouthparts beginning at the posterior end of the head and work anteriorly. The labium is the most posterior of the mouthparts (Fig 2, 4). It is a single unit composed of the two paired appendages of this, the labial, segment. It covers the other mouthparts and is the posterior wall of the preoral cavity. You can see that it is composed of a central region from which arise two short labial palps.
Grasp the free edge of the labium with your fine forceps and pull it ventrally and posteriorly to expose the remaining mouthparts. A large conic process on the inner surface of the labium fits into the mouth.
Maxilla
The two maxillae, which are not fused, are next (Fig 2). Each maxilla has a large antenna-like maxillary palp and a jagged, medial lacinia with many strong sclerotized teeth (Fig 4).
Mandibles
The mandibles are anterior to the maxillae and, like them, are not fused (Fig 2, 4). The mandibles lie on either side of the mouth and, along with the maxillae, form the lateral walls of the preoral cavity. Their most conspicuous feature is the large, amber, lateral, bladelike incisor used for shearing food into small pieces. Each mandible also bears a molar, used for grinding food. The black molar is medial and lies immediately beside the mouth.
Labrum
The anteriormost mouthpart, the labrum, is a small triangular green plate immediately ventral to the clypeus and dorsal to the mouth and mandibles (Fig 4). The mandibles are located beside and posterior to the labrum. The labrum is the anterior wall of the preoral cavity. Some zoologists believe the labrum to be derived from an anterior pair of appendages that have fused to form a relatively featureless plate. In support of this hypothesis the labrum is equipped with extrinsic muscles which move it with respect to the clypeus. It is innervated by the tritocerebrum suggesting it is the homolog of the crustacean second antenna.
Thorax
Legs
The legs should be studied now, after the head and prothorax have been removed from the remainder of the body.
Forelegs
Look at one of the forelegs. Each consists of the six articles of the typical insect leg (Fig 5). The cylindrical coxa is very large and appears to be part of the sternite of its segment. It fits snugly in a cylindrical recess in the prosternite and is easy to miss because it looks like a sternite and is hidden by the femur. If you grasp the leg and move it, you will see the coxa rotate on its long axis, thus demonstrating that it is not a sternite. The free edge of the coxa bears a fringe of white setae. The trochanter is a small triangular article at the distal end of a short process from the coxa. The femur is the long thick article extending distally from the trochanter. It is about the same size as the coxa. Following the femur is the tibia, which is also long. The foretibia has a conspicuous process extending distally from the distal end well past the point of articulation with the tarsus. This process makes the tibia the longest article in the foreleg.
Figure 5. Foreleg of Popillia. Beetle69L.gif
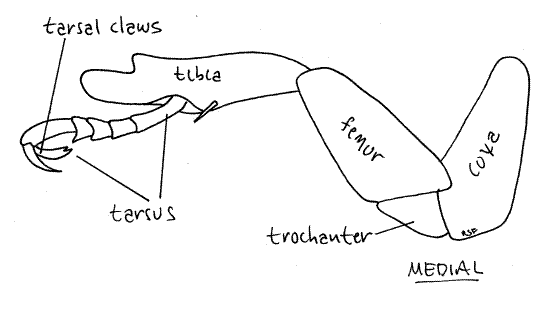
The tarsus is the last article of the insect leg but is divided into several smaller articles, the tarsomeres, arrayed in sequence. The foretarsus, which is the tarsus of the foreleg, consists of five tarsomeres. The distal tarsomere bears two long tarsal claws, one of which has one tooth and the other two. Together the tarsal claws plus any other distal accessories, are known as the pretarsus which is the sixth leg article.
Middle Leg
Now examine a midleg. It resembles the foreleg but is larger and its tibia does not have the distal process. Its cylindrical coxa fits neatly into a recess in the sternite. Find and name the articles of this appendage.
Hindleg
The hindleg is largest of the legs but it resembles the others in most respects (Fig 6). Its coxa differs, however, in being a flat plate that you may earlier have mistaken for a sternite (Fig 3). Grasp the coxa with your fine forceps and twist the leg off the metathorax. The actual extent and shape of the hindcoxa is now apparent. Both hindtarsal claws have a single point. Find and name the articles of this appendage.
Figure 6. Hindleg of Popillia. Beetle70L.gif
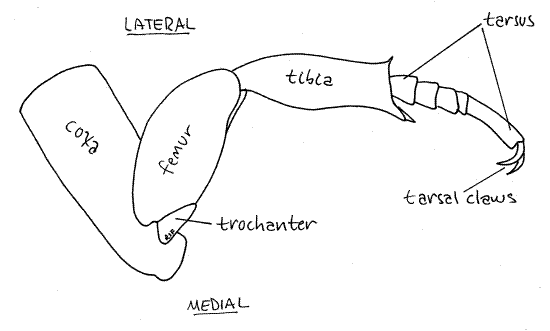
The number of tarsomeres is sometimes different in different legs and also varies with taxon. It can be an important taxonomic character used in the identification of insects. The number of tarsomeres in the three legs is indicated by the tarsal formula which is a series of three numbers such as "3,4,4". This means the foretarsus has three tarsomeres, the midtarsus has four, and the hindtarsus has four. What is the tarsal formula of Popillia japonica? _____, _____, _____
References
Borrer DJ, Triplehorn CA, Johnson NF. 1989. An introduction to the study of insects, 6 th ed. Saunders, Philadelphia. 875pp.
Ruppert EE, Fox RS, Barnes RB. 2004. Invertebrate Zoology, A functional evolutionary approach, 7 th ed. Brooks Cole Thomson, Belmont CA. 963 pp.
Supplies
Dissecting microscope
Living or preserved Japanese beetle
Anesthetic (carbon dioxide or chloroform) for living beetles
40 % isopropanol for preserved beetles
Fine forceps and teasing needles