Invertebrate Anatomy OnLine
Periplaneta americana ©
American Cockroach
30jun2006
Copyright 2004 by
Richard Fox
Lander University
Preface
This is an exercise from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises can be accessed by clicking on the links in the column to the left. A glossary and chapters on supplies and laboratory techniques are also available. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Arthropoda P, Mandibulata sP, Tracheata, Hexapoda SC, Insecta C, Dicondylia, Pterygota, Metapterygota, Neoptera, Blattaria O, Blattoidea SF, Blattidae F, Blattinae sF (Fig 16-15, 20-14, 20-15, 21-23)
Arthropoda
Arthropoda, by far the largest and most diverse animal taxon, includes chelicerates, insects, myriapods, and crustaceans as well as many extinct taxa. The body is segmented and primitively bears a pair of jointed appendages on each segment. The epidermis secretes a complex cuticular exoskeleton which must be molted to permit increase in size. Extant arthropods exhibit regional specialization in the structure and function of segments and appendages. The body is typically divided into a head and trunk, of which the trunk is often itself divided into thorax and abdomen.
The gut consists of foregut, midgut, and hindgut and extends the length of the body from anterior mouth to posterior anus. Foregut and hindgut are epidermal invaginations, being derived from the embryonic stomodeum and proctodeum respectively, and are lined by cuticle, as are all epidermal surfaces. The midgut is endodermal and is responsible for most enzyme secretion, hydrolysis, and absorption.
The coelom is reduced to small spaces associated with the gonads and kidney. The functional body cavity is a spacious hemocoel divided by a horizontal diaphragm into a dorsal pericardial sinus and a much larger perivisceral sinus. Sometimes there is a small ventral perineural sinus surrounding the ventral nerve cord.
The hemal system includes a dorsal, contractile, tubular, ostiate heart that pumps blood to and from the hemocoel. Excretory organs vary with taxon and include Malpighian tubules, saccate nephridia, and nephrocytes. Respiratory organs also vary with taxon and include many types of gills, book lungs, and tracheae.
The nervous system consists of a dorsal, anterior brain of two or three pairs of ganglia, circumenteric connectives, and a paired ventral nerve cord with segmental ganglia and segmental peripheral nerves. Various degrees of condensation and cephalization are found in different taxa.
Development is derived with centrolecithal eggs and superficial cleavage. There is frequently a larva although development is direct in many. Juveniles pass through a series of instars separated by molts until reaching the adult size and reproductive condition. At this time molting and growth may cease or continue, depending on taxon.
Mandibulata
Mandibulata includes arthropods in which the third head segment bears a pair of mandibles. As currently conceived this taxon includes myriapods, hexapods, and crustaceans. Appendages may be uni- or biramous and habitats include marine, freshwater, terrestrial, and aerial.
Tracheata
Myriapods and hexapods share tracheae and a single pair of antennae and are sister taxa in Tracheata. Crustaceans, which have gills and lack tracheae, are excluded and form the sister group.
Hexapoda
The body is divided into three tagmata; head, thorax, and abdomen. Appendages are uniramous and a single pair of antennae is present. Three pairs of legs and two pairs of wings are found on the thorax of most adults. Hexapod legs are uniramous although there is increasing evidence that they evolved from multiramous appendages of their ancestors. Gas exchange is accomplished by trachea. Excretory organs are Malpighian tubules and the end product of nitrogen metabolism is uric acid. There is relatively little cephalization of the nervous system. Insects are gonochoric with copulation and internal fertilization.
Insecta
Most hexapods are insects. A few hexapod taxa (orders) lack wings and have primitive mouthparts recessed into the head and belong to Entognatha, the sister taxon of Insecta. Insects have ectognath mouthparts and the adults (imagoes) of most taxa have wings.
Pterygota
Pterygotes are the winged insects. These insects are derived from a winged common ancestor. Adults of most taxa have wings although they have been lost in some.
Blattaria O
Roaches are fast running insects probably closely related to termites (Isoptera ). Some primitive taxa have endosymbiotic gut zooflagellates similar to those of termites. The tarsi are five-articulate. The oval body is dorsoventrally depressed and most the head is hidden from dorsal view by the large pronotum. The antennae are long and filamentous. Most taxa have wings, in at least one sex, but they are not always functional. Eggs are enclosed in a proteinaceous ootheca which may be gestated internally, brooded externally, or deposited without further maternal attention. Most cockroaches are tropical and several species are pests in or near buildings especially in warm climates. It is often found in homes and in my experience in the southeastern US, it is common in zoology laboratories, especially in those in older buildings.
Biology
Periplaneta americana
Periplaneta americana, the American cockroach, the largest widespread North American cockroach, reaches 44 mm in length. It is dark reddish-brown with a pale peripheral band around the pronotum. Adults have large wings and can fly, although they do not often do so. Nymphs, of course, are wingless. It is easily, albeit unintentionally and regretfully, spread by human commerce and is worldwide in warm climates and, in heated buildings, cosmopolitan. Periplaneta is nocturnal, negatively phototactic, and prefers dark warm, moist habitats. It is acutely sensitive to vibrations and is one of the world’s fastest running insects, as you know if you have tried to stomp one in the kitchen late at night. Periplaneta americana is one of several cockroaches found near (peridomestic) or in (domiciliary) human habitations. Such insects are referred to as synanthropic (= with man). Several of the most common North American synanthropic species are discussed below. Any of these species could be used for this exercise but other species of Periplaneta differ the least from this account and for our purposes are interchangeable with P. americana.
The life cycle consists of egg, numerous nymphal instars (6-14), and the adult, or imago. Blattarians are paurometabolous with nymphs closely resembling small adults except for the absence of wings. Paurometabolous insects have no pupa or metamorphosis. The eggs are laid in a tough, sclerotized protein case, the ootheca, normally containing 16 eggs. Females average one ootheca per month for most of the year but may exceed that average during periods of peak reproductive activity. Nymphs lack functional wings, of course, but wingpads appear in the 4 th instar and gradually increase in size with each molt. The life span is 2-3 years with only the last year being spent as an adult. Adults live for about a year.
This omnivorous, opportunistic, scavenging species feeds on almost anything organic, plant or animal, but prefer starches and sugars. Adults and nymphs have similar feeding niches.
Cockroach population density is controlled naturally by several species of parasitoid wasps including Evania and Aprostocetus. Both are egg parasitoids in which the female wasp oviposits in the roach ootheca where the wasp larvae hatch and feed on roach eggs. Evania is a distinctive, small, black wasp with a laterally flattened triangular abdomen that waves flag-like (hence the common name “ensign" wasp) as the adult walks. Adult Evania are about 1.5 cm in length. The female, with great difficulty, inserts one egg in each ootheca. The larva hatches and consumes all 16 roach eggs before achieving a length of 8 mm, pupating, and emerging from the now empty ootheca. Adult Aprostocetus are tiny, about 2 mm long. Both species are stingless and no threat to humans. Small wasps, especially those waving a flag, seen in roach-infested areas should be protected and encouraged, not killed. Attempts to control roach populations with pesticide sprays can be counterproductive because they indiscriminately kill parasitoids in addition to the roaches. Roach pellets (“roach hotels”), which are selective, are more effective.
Cockroaches are not biological vectors for human disease although they can serve as mechanical vectors simply by harboring infectious organisms such as Ascaris eggs, bacteria, or protozoan cysts on their body surfaces. The American cockroach is the host for the cystacanth stage of the rat intestinal acanthocephalan, Moniliformis moniliformis.
The Major Synanthropic Cockroaches
Seven species of cockroaches enjoy close synanthropic associations with humans and have been spread throughout the world by human migrations and commerce and are now serious pests. All these species are thought to have originated in Africa. Many other species are loosely associated with humans and their activities.
The American cockroach, Periplaneta americana , despite its specific and common names, is an exotic that was introduced to North America from Africa in the early 17 th century. It is the largest of the domiciliary species, reaching 44 mm in length. Both sexes have fully functional wings but rarely fly. The pronotum is bordered by a pale yellowish margin. It is easily confused with P. bunnea. The last article of the cercus of P. americana is over twice as long as wide (Fig 16) whereas that of P. brunnea is less than twice as long as wide (Fig 14). Periplaneta americana is the second most abundant cockroach pest in the United States.
Periplaneta fuliginosa (smoky brown cockroach) is an introduced peridomestic species. It is common in the southern United States mostly found out of doors or in crawl spaces or unheated buildings such as outhouses or garages. It also occurs in buildings in the south and can be found further north in heated buildings. In some localities it may account for almost 80% of cockroaches captured near homes. The color is uniformly dark brownish black and it lacks the pale pronotal margin of other synanthropic Periplaneta. Nor does it have pale areas on the forewings that distinguish P. australasiae. It reaches 35 mm in length, not quite as large as P. americana
Periplaneta brunnea (brown cockroach) is an African native spread by human travel and commerce. It is common in the southeastern United States, sometimes moreso than P. americana, but it is usually found outside. This species has a pale border around the pronotum as do most Periplaneta but it lacks the pale areas on the forewings characteristic of P. australasiae. It reaches 37 mm in length. Both sexes are winged and can fly.
Periplaneta australasiae (Australian cockroach) is almost as serious a domestic pest as is P. americana. It probably arose in Africa and spread throughout the tropics and subtropics and is now circumtropical. It is the most common domiciliary cockroach in Florida and although it occurs farther north it is intolerant of the cooler temperatures tolerated by P. Americana. It is similar to P. americana but slightly smaller, reaching 35 mm in length. It is reddish brown, like P. Americana. Unlike our other Periplaneta species, its forewings have a pale yellowish band on the proximal anterior edge. Both sexes have wings and fly. The specific epithet “australasiae” means “south Asia”, not Australia.
Blatta orientalis (oriental cockroach, black beetle, water bug) probably originated in North Africa and has become a common pest in the southern, midwestern, and northwestern United States but is known as far north as southern Canada, the Netherlands, and Britain. It prefers cool, moist habitats. Males are winged, females have tiny vestigial wings, and neither is capable of flight. Individuals reach 27 mm in length. Its color is dark, almost black, and it prefers moist habitats. Blatta lacks an arolium on its tarsi and cannot climb smooth surfaces. The common name “water bug” derives from its preference for wet areas.
The German cockroach, Blatella germanica , originated in Africa and traveled first to Europe and from there to North America. It is the most abundant domiciliary cockroach in the United States and is an important pest. At 15 mm, it is much smaller than the Periplaneta species. It is pale brown with two dark longitudinal bands bordering the pronotum.
Supella longipalpa (brown banded cockroach) is assumed to have originated in Africa. It is always found in buildings (except in Africa). It was originally introduced into the Florida and spread north hitchhiking on fruit shipments and in the luggage and automobiles of tourists returning north after a vacation. It is now found throughout the contiguous United States. This is a small roach, reaching only 14 mm, and similar in size and general appearance to Blatella germanica. The forewings cover the abdomen of the male but those of females are shorter and the posterior abdomen is left uncovered. Two pale bands extend transversely across the dark abdomen but these may be difficult to see with the wings folded at rest. They are obvious in the wingless nymphs. Supella lacks the longitudinal stripes on the pronotum characteristic of Blatella. The lateral margins of the pronotum of Supella are transparent.
Laboratory Specimens
Periplaneta is available at modest cost, alive or preserved, from biological supply companies. It can also be captured locally in homes or university buildings. Look for it in warm, moist, dark places such as basements, drawers, wood piles, sewers, sunken water meter enclosures, and compost piles. In the home it can be found wherever there is food, including the usual debris on the kitchen floor and under the refrigerator, on unwashed dishes left overnight in the sink, and even in your library where it eats the bindings of your favorite books. Female Periplaneta are distinguished from other genera by having a divided 7 th sternite.
Specimens may be dissected alive and anesthetized, recently sacrificed and unpreserved, or preserved. Recently sacrificed is preferable unless you anticipate making physiological observations, in which case the specimens should be alive and anesthetized. Living specimens may be sacrificed in a covered dish with a cotton ball dampened with ethyl acetate. Living specimens should be anesthetized using chloroform, ether, or carbon dioxide. Sacrificed specimens should be immersed in water or 80% ethanol (or 40% isopropanol) in a small dissecting pan.
If both external and internal anatomy are to be studied it is advantageous to have separate specimens for these two undertakings. The study of external anatomy is likely to damage the specimen to such an extent that it will not be useful for studying internal anatomy. Students should have access to both sexes, either their own specimens or by sharing with a lab partner.
Anatomy
External Anatomy
Species of Periplaneta are elongate, oval in outline and strongly dorso-ventrally depressed, or flattened. The body is divided into the three tagmata characteristic of insects, i.e. head, thorax, and abdomen (Fig 21-1C). The head is inconspicuous in dorsal view but the pronotum of the thorax is very large. The remainder of the thorax and abdomen are hidden by the two pairs of wings. Six pairs of similar, strong, spiny legs are present.
Study a recently sacrificed or preserved cockroach with the dissecting microscope. The specimen may be dry or submersed in liquid.
Figure 1. Dorsal view of a female smoky brown cockroach, Periplaneta fuliginosa, from Greenwood, South Carolina. Blatt37L.gif
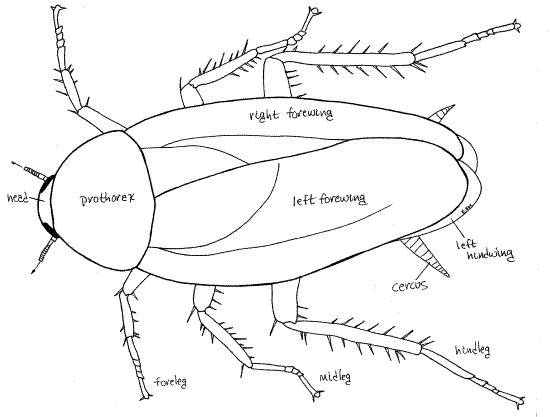
Head
Little of the head can be seen in dorsal view (Fig 1). Position the specimen with the ventral surface up and examine the head with the dissecting microscope. The head is flattened dorsoventrally and is more or less pear-shaped in outline with the mouthparts extending posteriorly from the narrow end of the pear. The head is normally carried with its flat front surface, the face, held horizontally facing the substratum (Fig 2). In this position the mouth opens posteriorly, the mandibles project posteriorly, and the palps touch the coxae of the forelegs. When feeding, the flattened face is held vertically with the mouthparts located ventrally, toward the substratum. The flattened face is the anterior aspect of the head even though it is oriented ventrally. Similarly, the thin edge of the broad end of the pear is dorsal, even though it is normally held in an anterior position. When in the feeding position the face is held anteriorly and the thin edge is dorsal.
Cockroaches have typical unspecialized mouthparts and provide a good example of the primitive insect mouthparts adapted for biting and chewing (Fig 21-1A,B). The head can be retracted beneath the prothorax.
Figure 2. Anterior view of the head of Periplaneta fuliginosa. Blatt38L.gif
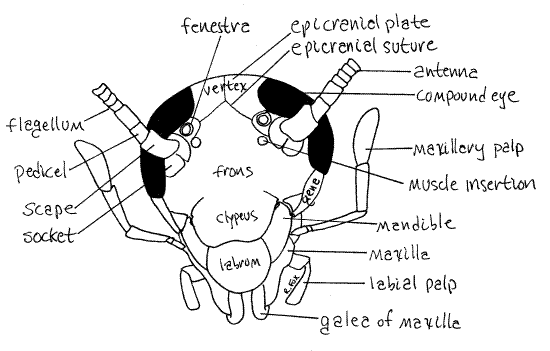
Head Capsule
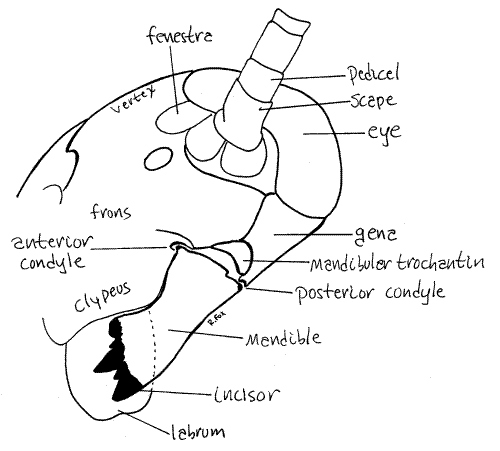
The head consists of a complex of fused sclerites, which constitute the epicranium, or head capsule, associated with unfused sclerites and appendages. This hard sclerotized capsule encloses the mouthpart muscles, brain, and anteriormost region of the gut and provides sites of attachment for the mouthparts and antennae.
The head capsule is divided into several regions. Locate the large compound eyes on the dorsal edge of the head (Fig 1, 2). The antennae arise on the face immediately ventral to the eyes. These are useful as convenient, easily recognized landmarks.
The epicranial suture marks the junction of three fused sclerites that form the face. In Periplaneta this suture is usually visible as a faint, yellow, Y-shaped line on the face (Fig 2) but sometimes it cannot be found. The two arms of the "Y" begin at the antennal sockets and converge between the two eyes. From the convergence the single stem of the "Y" extends dorsally between the eyes over the top of the head. It divides the top of the head into a pair of sclerites, the epicranial plates (Fig 2). The part of the capsule between the two arms of the “Y” is thefrons, or front, of the capsule (Fig 2). The frons is an unpaired sclerite.
Two white circular areas between the antennae and compound eyes, at the ends of the arms of the epicranial suture, are the fenestrae and are the vestiges of the ocelli of other insects (Fig 21-1A). Two additional pale circles on the frons, ventral and a little medial to the fenestrae, mark the sites of muscle insertions inside the capsule.
The vertex, which is a region, not a sclerite, occupies the dorsum of the capsule between the two eyes. It is divided in two by the stem of the epicranial suture (Fig 2) and includes the two epicranial plates. In some insects the posteriormost region of the vertex may be a distinct sclerite referred to as the occiput.
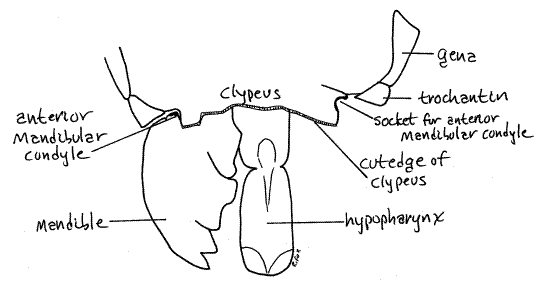
The clypeus (KLIP ee us) is an unpaired sclerite on the face (Fig 2, 21-1A). It lies ventral to the frons and the two are joined along a transverse, indistinguishable suture. The clypeus is not movable.
A third unpaired sclerite, the labrum, is attached along a transverse, movable articulation with the ventral edge of the clypeus (Fig 2). The labrum is equipped with muscles and is movable. It forms the anterior wall of the preoral cavity into which the mouth opens. The labrum is usually considered to be derived from a sclerite of one of the head segments but some entomologists believe it to be homologous to a pair of fused appendages, similar to the labium in this regard.
The sides, or cheeks, of the epicranium are formed by the paired genae. In the cockroach these form the thin lateral edges of the head ventral and posterior to the eyes. The genae are formed of several indistinguishably fused sclerites.
Posteriorly the head capsule is penetrated by a large circular opening, the foramen magnum through which pass the gut, salivary ducts, and nerve cord. The foramen is bordered by the vertex dorsally and the genae laterally. The soft, cervix, or neck, extends posteriorly from the foramen. Because of the attachment of soft tissue of the cervix to the head capsule, the foramen is not visible externally. Ventrally slender, transverse cervical sclerites are embedded in the otherwise soft integument of the cervix.
Antenna
The anteriormost head appendages are the two antennae. These arise from a socket ventral to the compound eyes. The first article of the antenna is the scape and the second is thepedicel (Fig 2). From the pedicel arises a long, whiplike, sensory flagellum of 75-90 articles. The antennae are equipped with mechanoreceptors but apparently not with chemoreceptors.
Preoral Cavity
Ventrally the mouthparts surround and enclose a chamber, the preoral cavity, from which the mouth opens (Fig 21-7). As its name suggests, the preoral cavity precedes the mouth and is not considered to be part of the gut, although both mechanical and chemical digestion begin there.
With the specimen positioned with its ventral side up, grasp the labrum and mandibles with a pair of fine forceps and pull the head into the feeding position (i.e. with the anterior face anterior, and the mouthparts ventral). Lift the labrum and look beneath it using a second pair of forceps or a fine needle to move structures aside as needed.
The labrum, which you are currently holding aside, is the anterior wall of the cavity. The heavy, toothed mandibles and softer maxillae are its side walls and the labium is its posterior wall. A soft, tonguelike, mostly unsclerotized outgrowth of the ventral body wall, the hypopharynx, extends into the preoral cavity immediately posterior to the mouth. Move the mandibles and maxillae aside and find the mouth opening dorsally from the preoral cavity immediately anterior to the hypopharynx.
Mandible
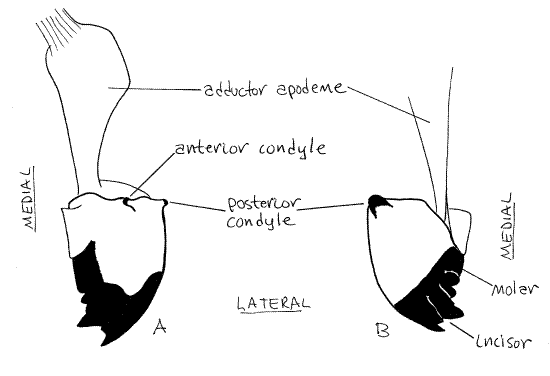
The two mandibles lie beside the mouth and form the lateral walls of the preoral cavity (Fig 2, 21-1A, B). They are derived from a pair of segmental appendages. Each mandible consists of a single article bearing a sclerotized, distal, cutting surface. The cutting edge of the mandible consists of a lateral shearing incisor with three or four sharp denticles and a more medial grinding molar (Fig 3, 21-1A).
The mandible is largely hidden from view but can be glimpsed on the side of the head ventral to the gena and lateral to the clypeus (Fig 2, 4). With forceps carefully lift the labrum and peer beneath it to get a better view of the mandible. Its dark, heavily sclerotized denticles are visible in this view.
The mandible articulates with the head capsule by two ball and socket-like condyles, one anterior and one posterior (Fig 3, 4). The anterior condyle can be seen at the dorsolateral corner of the clypeus, on the face. The posterior condyle articulates with the ventral corner of the gena and can be seen on the side of the head capsule. Its plane of motion is transverse, perpendicular to the long axis of the body. Movement in this plane brings the two mandibles together on the midline. In contrast, vertebrate mandibles oppose each other by moving in the vertical plane. Observe the mandible with the labrum moved aside and imagine the motion of the two mandibles with respect to each other. Grasp the mandibles with forceps and move them in their preferred plane of motion. If desired, one of the mandibles can be removed for closer examination.
Figure 3. The left mandible. A. Anterior view. B. Posterior view. Blatt39L.gif
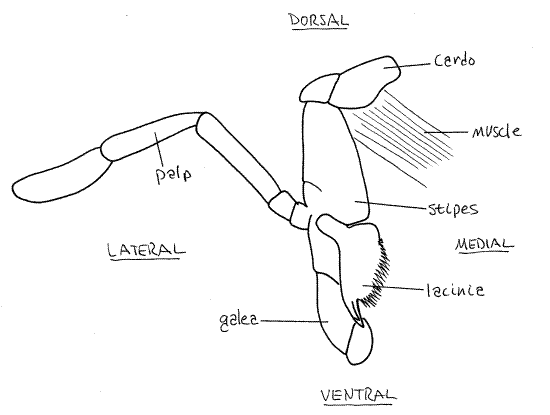
Maxilla
Look at the head from the side and find the maxilla (Fig 2, 21-1A,B). Like the mandibles, the two maxillae are paired appendages lying beside the preoral cavity. The maxilla can be recognized by its large 5-articulate palp (the labium also has a palp but it is much smaller and has three articles). Each maxilla consists of a proximal cardo that articulate with the head capsule and a distal stipes from which arise several processes.
Figure 4. Oblique view of the head from the left showing the articulation of the mandible with the head capsule of P. fuliginosa. The labrum has been drawn as if transparent to reveal the left mandible beneath it. The right mandible has been omitted for clarity. Blatt40L.gif
The cardo in a short transverse partly sclerotized article extending laterally from the head capsule (Fig 5). It is inconspicuous because it is hidden by the larger stipes. The stipes is easily seen when looking at the head from the side. It is elongate and articulates with the cardo proximally, beside the ventral edge of the eye. Distally three processes arise from the stipes.
Most obvious of these processes is the maxillary palp consisting of five articles. The palp is sensory with chemo- and mechanorecepotors used to evaluate the suitability of potential food. From the distal end of the stipes arise a lightly sclerotized, lateral galea and median, sclerotized, bladelike lacinia, which bears denticles distally. The lacinia assists the mandible in cutting food into smaller particles. The lacinia is completely hidden by the soft bulbous galea. The galea acts as a sheath to cover the lacinia and because of it, the lacinia is not visible until the galea is moved aside. You may remove one of the maxilla for closer study if desired.
Labium
The insect labium, also known as the second maxilla, is formed by the fusion of the posteriormost pair of head appendages. The labium is the posterior wall of the preoral cavity. It is best viewed by lifting the head to reveal its posterior surface. The labium consists of a large, platelike, proximal submentum (Fig 6, 21-1A,B). Articulated with it is the similarly platelike, but smaller, mentum. Submentum and mentum are the fused regions of the ancestral appendages. Arising from the distal edge of the mentum is a pair of prementa, the homologs of the original two appendages. Each prementum bears a distal median glossa with a lateral paraglossa beside it. The glossae and paraglossae are together known as the ligula and function in the manipulation of food. A 3-articulate labial palp arises laterally from the side of each prementum. The palp is chemo- and mechanosensory. You may remove the labium and examine it with higher magnification if you wish.
Figure 5. Posterior view of the left maxilla of P. fuliginosa with the galea moved slightly to reveal the lacinia. Blatt41L.gif
Hypopharynx
The hypopharynx, or tongue, is a long, process protruding from the ventral wall of the head into the preoral cavity (Fig 7, 21-7). It is not a segmental appendage being instead a fold of the body wall. It can be seen by looking into the preoral cavity, either ventrally by spreading the mouthparts aside, or posteriorly by lifting (or removing) the labium. The hypopharynx divides the preoral cavity into an anterior cibarium and a posterior salivarium.
Thorax
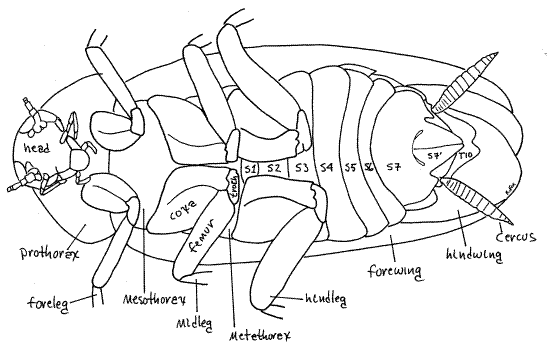
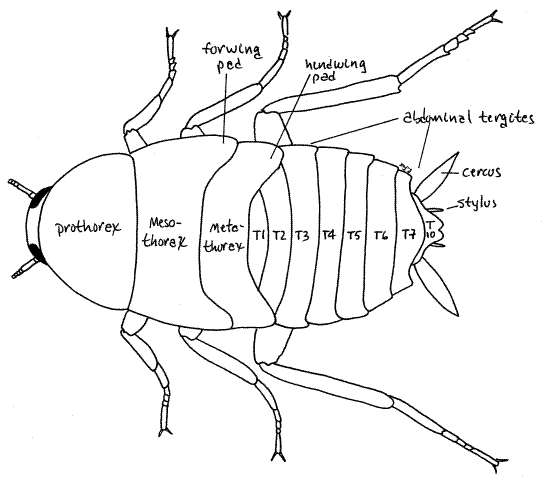
The thorax is the tagma specialized for locomotion and as such bears three pairs of legs and, in adults, two pairs of wings and houses the muscles to operate them (Fig 16-2). It is the middle tagma of the body and consists of the anterior prothorax, middle mesothorax, and posterior metathorax, of which the prothorax is by far the largest and the only one visible in dorsal view. In ventral view the prothorax, mesothorax, and metathorax are all visible and easily recognized by virtue of the pair of walking legs carried by each segment.
Figure 6. Posterior view of the labium. Blatt42L.gif
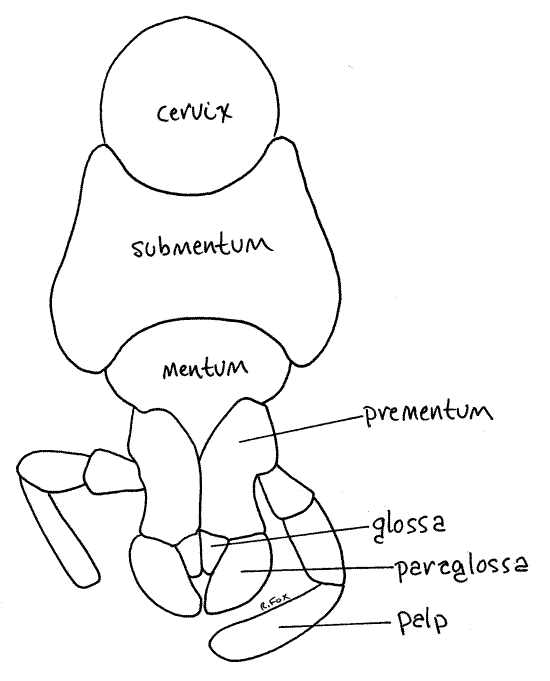
Figure 7. Anterior view of the hypopharynx of P. fuliginosa. The clypeus, labrum, left mandible, and labium have been removed for clarity. Blatt43L.gif
Legs
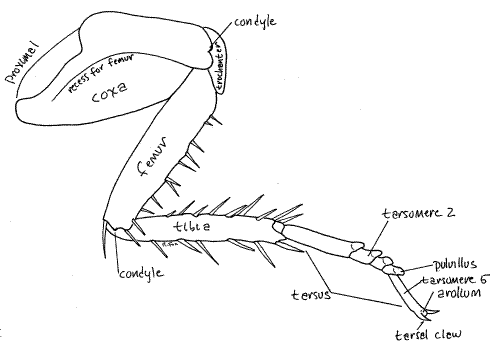
The large powerful legs are responsible for the cursorial competency for which cockroaches are renown. Three pairs are present, of course, one on each thoracic segment (Fig 8). The three pairs are similar but increase in size from anterior to posterior. Each consists of a large, flattened, proximal coxa, a small trochanter, a long femur, a tibia, and a long tarsus (Fig 9, 21-1E). The femur and tibia bear strong spines. The tarsus is a series of 5-articulate tarsomeres. Tarsomeres 1-4 each bear a posterior pad-like adhesive pulvillus. Tarsomere 5 ends with a pair of tarsal claws beside a pad-like arolium. This distal arrangement of claws and arolium is sometimes referred to as the pretarsus. The arolium is an adaptation for clinging to smooth surfaces and makes it possible for Periplaneta to climb smooth walls.
Figure 8. Ventral view of a female P. fuliginosa. The distal articles of the legs have been removed for clarity. S = sternite, T = tergite. Blatt44L.gif
When flexed, the femur fits neatly into a recess in the surface of the coxa (Fig 8). The articulations between coxa and trochanter and between femur and tibia are dicondylic joints which restrict movement to a single plane (as, you may recall, does the dicondylic joint between the mandible and head capsule). The coxa-body articulation is monocondlyic and permits movement in a variety of planes as is the tibia-tarsus articulation. Dicondylic joints are functionally similar to the hinge joints in your knee and elbow whereas monocondylic joints function like ball and socket joints in your shoulder and hip.
Wings
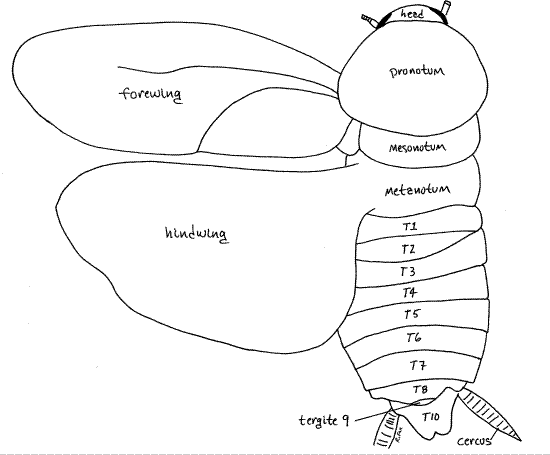
The meso- and metathorax of adults of both sexes of Periplaneta each bear a pair of large wings. The anterior mesothoracic wings, or forewings (= wing covers, often known as tegmina in orthopterans and blattarians), are obvious in dorsal view whereas the equally large metathoracic wings (hindwings) are almost entirely hidden by the forewings (Fig 10). The heavier, parchment-like forewings protect the more delicate hindwings. The hindwings are membranous and fold in pleats when at rest under the forewings. The folded wings completely cover the dorsal surface except for the head and pronotum.
Fig 9. Ventral view of the right midleg of P. fuliginosa. Blatt45L.gif
Use forceps and your fingers to extend the wings and examine their shape and texture. Note the way in which the hindwing folds when at rest.
Cockroach nymphs, like immatures of other insects, lack wings, but like other pauro- and hemimetabolous insects, develop wing pads in older instars (Fig 11). The wing pads are transformed to functional wings by the last molt.
Thoracic Segments
The thorax consists of three segments but your view is obscured dorsally by the wings and ventrally by the walking legs, especially their coxae. Each segment has the expected complement of sclerites consisting of dorsal tergite (or notum), lateral pleurites, and ventral sternites.
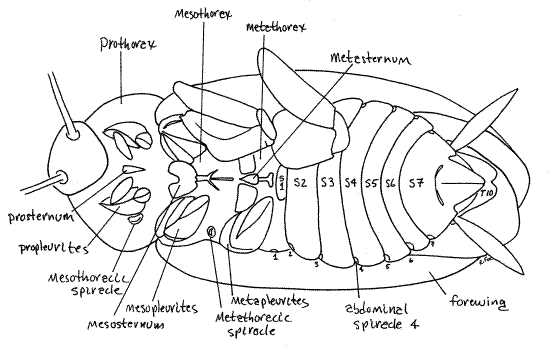
The ventral surface of the thorax consists of large expanses of unsclerotized, flexible, white exoskeleton in which are embedded myriad hard, golden brown, sclerites. Anchor the specimen on its back with a #1 stainless steel insect pin through the margin of the pronotum. Move the legs aside as needed to reveal the ventral surface of the thorax. The proximal end of the coxa of each thoracic limb articulates with a complex of sclerotized pleurites (Fig 12, 21-1D). On the midline, between the right and left pleurites, lies a series of thoracic sternites. Theprosternum is a small, triangular, median, unpaired sclerite in the center of the prothorax, between the clusters of prothoracic pleurites. The mesosternum is more complicated than the prosternum and consists of several sclerites including a large bilobed anterior plate followed by two slender posterior sclerites. The metasternum is similar to the mesosternum except that the bilobed plate is completely divided into a pair of plates.
Figure 10. Dorsal view of a female P. fuliginosa with the wings removed or extended to reveal the dorsal surface of the thorax and abdomen. The abdomen has been stretched slightly to reveal tergites 8 and 9 which are normally hidden under tergite 7. blatt46L.gif
The thorax is equipped with two pairs of spiracles which open into the tracheal respiratory system. The spiracles are lateral and belong to the mesothorax and metathorax although they have migrated anteriorly (Fig 12). Consequently, the mesothoracic spiracle is on the posterior prothorax and the metathoracic spiracle is in the membrane between the mesothorax and metathorax.
Study the dorsal thorax by moving the wings aside. The pronotum is not obscured by wings and is always easily seen. It forms a large shield behind the head and may be mistaken for the head by the uninitiated. It is heavily sclerotized and dark in color with a pale border. The mesonotum (Fig 10) is a transverse, lightly sclerotized plate covering the dorsal surface of the mesothorax. The metanotum is similar and covers the mesothorax.
Abdomen
The abdomen is the largest of the three tagmata and contains most of the viscera. Its segmentation is conspicuous both dorsally and ventrally once the wings have been removed. The abdomen consists of 11 segments of which the 11 th is reduced and fused with the 10 th so that a maximum of ten segments are distinct and visible (Fig 21-1F). Further, some of the segments, especially in females, are highly modified or in unexpected locations and are not immediately visible and countable.
Figure 11. A late instar, 24 mm Periplaneta nymph. Blatt47L.gif
The first seven segments, known as the pregenital, or visceral, segments are similar, unspecialized, visible and countable. Segments 9 in males and 8-9 in females are the genital segments and are modified for reproductive functions. Segment 10 (and the vestigial 11) are postgenital and are also modified.
Dorsally each segment is covered by a sclerotized tergite and, similarly, is covered ventrally by a sclerotized sternite. Consecutive sternites articulate with each other by unsclerotizedarticular membranes (Fig 15). Laterally, tergites and sternites are connected by unsclerotized flexible pleura.
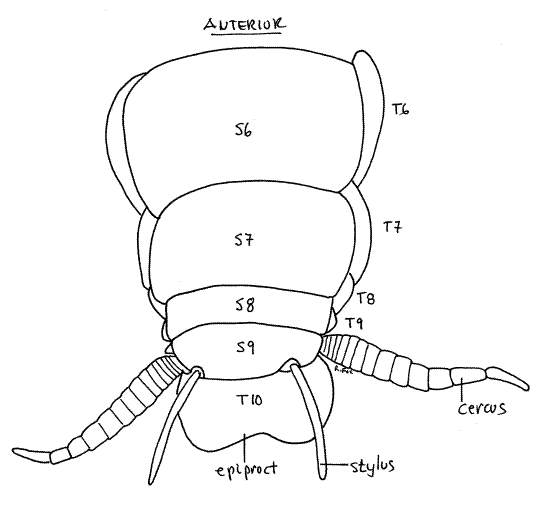
Examine the dorsum of the abdomen and find the abdominal tergites (Fig 10). Those of segments 1-7 are similar in size and easily distinguished but tergite 8 is smaller and tergite 9smaller still. These two tergites are usually telescoped under tergite 7 and may be difficult to see until you lift tergite 7. Tergite 10 is a large posteriorly bilobed plate overhanging the anus (Fig 10). Tergite 11 is fused with tergite 10. The bilobed posterior extension of tergite 10 is sometimes referred to as the epiproct, which is homologous to the tergite of segment 11 (Fig 10, 21-1F).
Study the ventral surface of the abdomen (Fig 12, H). Sternite 1 is a small sclerite lying on the midline between the hindleg coxae. Sternites 2-6 are elongate, more or less rectangular, sclerotized plates covering the ventral surface of the abdomen. Sternites 7-9 of males resemble sternites 2-6 but the posterior sternites of females are very different. In females the posteriormost visible sternite is the large, highly modified sternite 7 (Fig 12, 13). Sternites 8, 9 are internalized under tergite 7 and cannot be seen without dissection.
Figure 12. Ventral view of a female P. fuliginosa with the right legs removed to reveal the ventral body surface and its sclerites. The position of the abdominal spiracles under the overhang of the preceding sternite is indicated. Blatt48L.gif
The anus is on segment 11 and lies ventral to the epiproct (tergite 11). It is flanked by a pair of triangular sclerites, the paraprocts (Fig 14, 21-11B, 21-12B,C) which are modifications of sternite 11. Sternite 10 is vestigial.
Most abdominal segments lack appendages but segment 10 bears a pair of large, segmented cerci, which are visible in both dorsal and ventral views (Fig 8, 13, 21-1F). Embryologically the cerci arise as the appendages of the 11 segment but in adults are attached to segment 10 since segment 11 has fused with segment 10. The cercus, although segmented, does not contain intrinsic muscles. Extrinsic muscles, however, extend from the cercus to the abdomen so it is capable of motion. Cerci bear vibration-sensitive hair sensillae sensitive to air movements, including sound, and perhaps to ground vibrations.
Eight pairs of spiracles are present in the abdomen and are much smaller than the thoracic spiracles. The abdominal spiracles are located laterally on the anterior-dorsal corner of the pleura of the first eight abdominal segments (Fig 12, 14, 21-1D,F). Lift the free posterior edge of a sternite and look at the antero-lateral corner of the following tergite to see the spiracle. It will be a small white oval. Abdominal spiracle 1 is on the lateral edge of tergite 1 rather than at the corner (Fig 12).
Genital Segments
Female
Study the dorsum of the posterior abdomen of a female. The tergites of the genital segments (abdominal segments 8, 9) are reduced and their tergites are largely hidden by the overhang of tergite 7 (Fig 10). Tergite 10 (fused with tergite 11) is a large, thin, fan-like plate extending posterior to the genital segments. It overhangs the anus and genital pouch, which will be described shortly. In P. americana tergite 10 is cleft posteriorly (Fig 10).
Figure 13. Ventral view of the posterior end of the abdomen of a female of P. fuliginosa. Blatt49L.gif
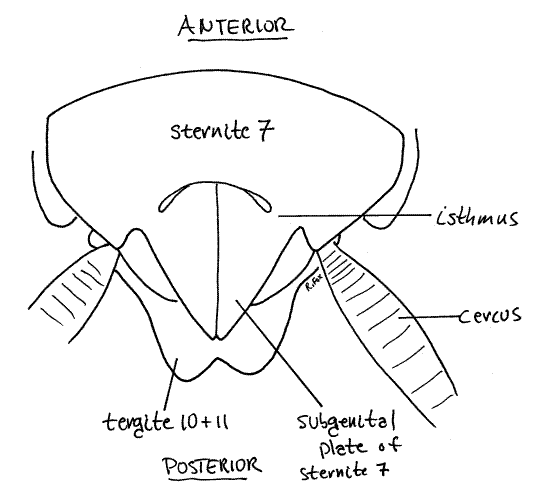
Look at the ventral surface of the abdomen (Fig 13). Sternites 8 and 9 cannot be seen externally. Sternite 7 (= hypogynum) is much larger and more heavily sclerotized than other sternites and posses a pair of clamshell-like posterior valves, the subgenital plates (also referred to variously as sternites 7 ¢ or 7a). Although at first the plates may appear to be independent of sternite 7, they are in fact continuous with it and connected by a short isthmus (Fig 13, 14). The isthmus is flexible and the plates can move apart during copulation or to permit extrusion of the ootheca as it is formed.
Hold the specimen upright and focus on the posterior end. This is much easier if you first separate the posterior abdomen from the remainder of the body (Do not remove the posterior abdomen if you plan to use this specimen later for study of internal anatomy). Make the separation between segment 6 and 7. You can now stand the posterior abdomen upright on the wax of the dissecting pan and you will not have to hold it in position. Focus on the posterior end of the abdomen (Fig 14).
Figure 14. Posterior view of the abdomen of a female P. fuliginosa. Arrows indicate movement of the subgenital plates. Blatt50L.gif
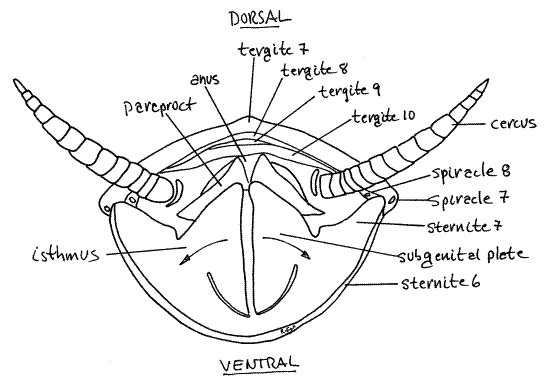
Tergites 7-10 dorsally and sternite 7 ventrally enclose a large space. The space is divided into dorsal and ventral regions by two lateral sclerites, the paraprocts (Fig 14, 21-11B). The anus opens into the dorsal chamber whereas the oviduct and seminal receptacle open into the ventral chamber, which is the genital pouch (Fig 15). With forceps pull the paraprocts aside to reveal the anus on segment 10-11 under the epiproct.
The genital pouch is formed by invagination of the exoskeleton of the sternum which brings sternites 8 and 9 deep into the pouch (Fig 15). This is the reason you could not see these sternites externally. With forceps and needles as needed, pull the subgenital plates of sternite 7 laterally and ventrally to reveal the genital pouch (= gynatrium). The genital pouch receives the spermatophore from the male during copulation and later the ootheca is molded in it.
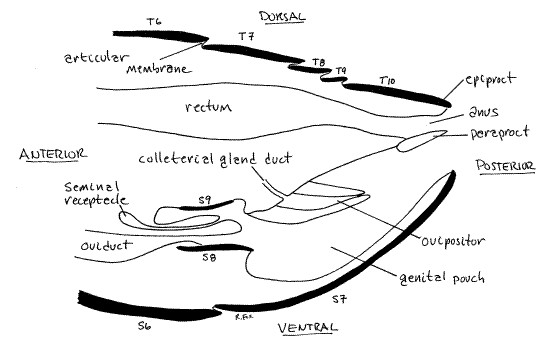
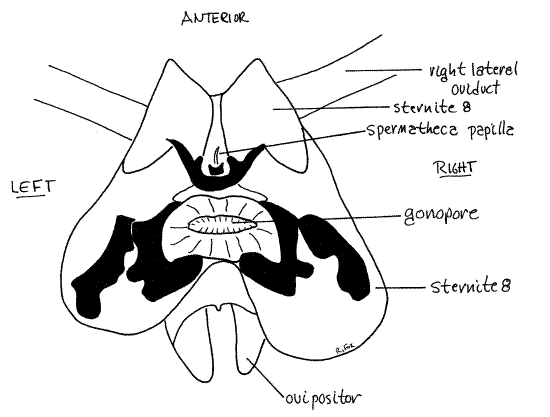
The most conspicuous feature in the genital pouch is the ovipositor (Fig 15). It is a median process arising from the roof of the genital pouch and consists of three pairs of cuticularizedvalvulae (= gonapophyses). The valvulae are derived from the paired appendages of segments 8 and 9. The ovipositor guides eggs from the oviduct to the forming ootheca. The male external genitalia grip the ovipositor during copulation.
Sternite 8 can be seen by lifting the ovipositor dorsally and looking under it Fig 15, 23). Sternite 8 is a large sclerotized plate penetrated by the female gonopore opening from the common gonoduct. Sternite 9 is at the base of the ovipositor.
Figure 15. Sagittal section of the posterior abdomen of a generalized female cockroach. S = sternite, T = tergite. Redrawn from Cornwell (1968) after McKittrick (1964). Blatt51L.gif
Male
The dorsum of the male posterior abdomen is similar to that of the female. Tergite 7 is large and tergites 8 and 9 are much smaller and obscured by the overhang of 7 (Fig 16). Tergite 10 is a large, posteriorly cleft plate fused with tergite 11 to form the epiproct.
Figure 16. Dorsal view of the posterior abdomen of P. americana. T = tergite. Blatt52L.gif
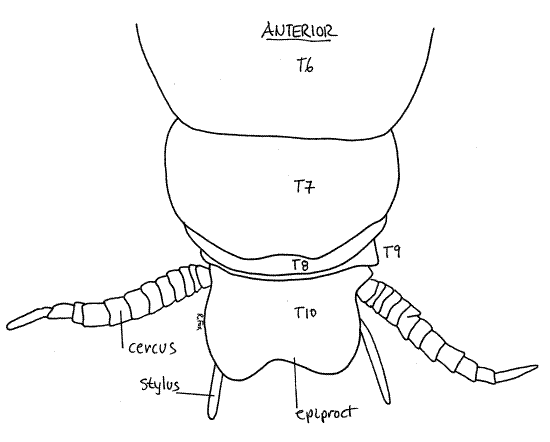
The male venter is very different from that of females. Sternites 8, and 9 are unremarkable and unmodified from the condition of the more anterior sternites and visible externally (Fig 17). Sternite 9 is the posteriormost sternite and its posterior border bears a slender stylus on each side. Styli are absent in mature females but are present in nymphs of both sexes (Fig 21-12B).
A pair of triangular paraprocts is located immediately ventral to tergite 10-11 and lateral to the anus (Fig 14). Between the paraprocts and sternite 9 is a large genital pouch in which are housed the male external genitalia.
Figure 17. Ventral view of the posterior abdomen of a male P. americana. Blatt53L.gif
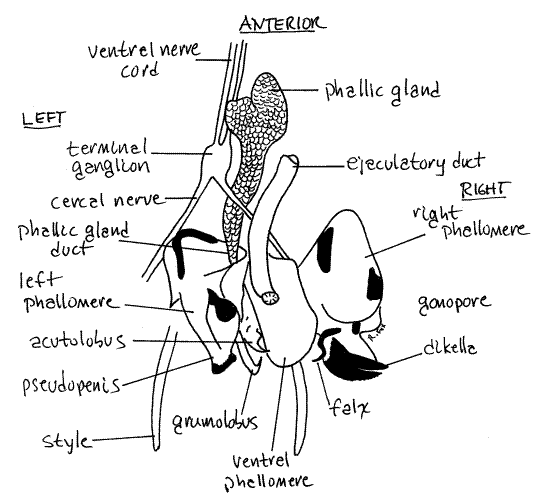
Anchor the posterior abdomen, ventral side up, with a pair of pins passing through the lateral tergites of segment 7. Lift or remove sternites 8 and 9 to reveal the genital pouch and its contents (Fig 18, 24). The male external genitalia comprise the several complex sclerites and soft tissues of three phallomeres (= phallic lobes, = gonapophyses), surrounding the male gonopore. The phallomeres are responsible for moving the subgenital plates aside to open the female’s genital pouch and for holding the female genitalia during copulation. Muscles arising on segment 9 extend to the phallomeres and operate them. Phallomeres are the appendages of segment 9.
Viewed from the venter, the ventral phallomere (Fig 18) is the most obvious of the three and can be seen immediately under sternite 9 (which has been removed or lifted). It is ventral to the gonopore and is by far the simplest of the three lobes. Its ventral surface is a slightly cupped sclerotized plate but it is membranous dorsally. The membranous ejaculatory duct ends at the gonopore on the dorsal surface of its base. Because it bears the gonopore, it is sometimes known as the penis.
The left phallomere is left of the ventral lobe and a little dorsal to it (Fig 18). It is a complex structure consisting of many sclerotized pieces including a hooked lobe (= grumolobus, titillator), prickly lobe (= acantholobus), a sharp stylet (= acutolobus, asperate lobe), and a pseudopenis. These are used during copulation to hold the valvulae of the female ovipositor so the spermatophore can be deposited in the correct location (at the opening of the seminal receptacle).
Figure 18. Ventral view of the posterior abdominal segments of a male P. americana. Sternites 8-9 have been removed and the phallomeres rearranged slightly for clarity. Some membranes and connective tissue have been removed. S = sternite, T = tergite. Blatt54L.gif
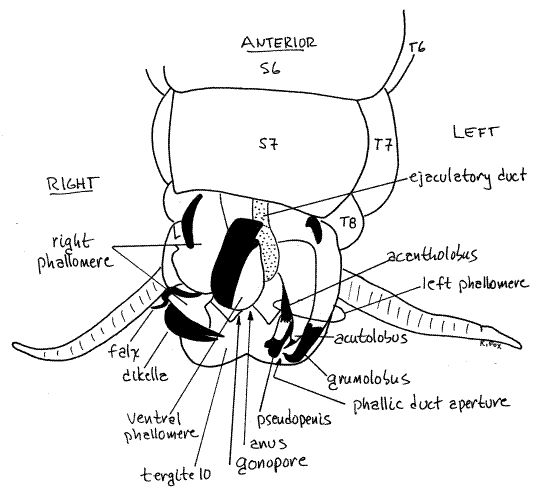
The right phallomere is to the right of the ventral phallomere and gonopore and is farther dorsal than either of the other two lobes. It is less complicated than the left lobe but also has easily recognized sclerotized parts. These include a piece that looks like a crayfish cheliped (= dikella, serrate lobe) and slender hooked piece (= falx).
Internal Anatomy
The cricket, Acheta, is recommended as an alternative for the study of internal anatomy in preference to the cockroach. Even though it is smaller, it is easier to dissect and its internal structures are easier to demonstrate.
Begin the study of cockroach internal anatomy with a fresh, undamaged specimen if possible. The dissection will be made from the dorsal side so you must first remove the wings by cutting their attachments with a pair of fine scissors. The dissection is facilitated if the legs are removed by cutting across their trochanters. Organ systems will be considered in order of their appearance in dorsal dissection.
" Place the specimen in a small dissecting pan of water or alcohol so it is completely immersed. Insert the blade of a pair of fine scissors under the posterior overhang of the right side of tergite 7 about 1 mm from the lateral margin of the tergite. Insert the blade only as deep as is necessary to penetrate the exoskeleton and be careful you do not damage internal organs with deep cuts. Cut anteriorly along the right side of the tergites all the way to the anterior end of the pronotum (Fig 19). Cut transversely across the anterior margin of the pronotum, just posterior to the head, and upon reaching the left side, change directions and cut posteriorly along the left side all the way back to tergite 7. Make a transverse cut through the exoskeleton across the posterior border of tergite 7. You have now cut all of the way around the dorsum.
Figure 19. Dorsal view of a male P. americana. Thoracic nota and abdominal terga 1-7 have been removed. T = tergite. Blatt55L.gif
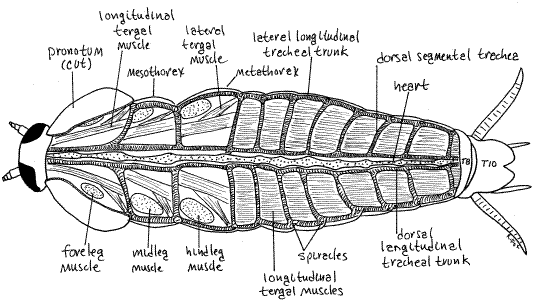
Anchor the specimen to the wax of the dissecting pan with a #1 stainless steel insect pin through the left side of one of the thoracic segments, lateral to your incision. Insert the pin at a 45 ° angle. Remove the abdominal and thoracic tergites beginning with tergite 7. Lift each tergite and remove it without removing any of the underlying soft tissues. The tergites are transparent and thin whereas the underlying tissues are opaque. These tissues include muscles, heart, and tracheae. Do not remove the opaque tissue at this time.
In the abdomen the tergal muscles form a broad thin sheet of longitudinal muscle fibers (Fig 19). In the thorax the tergal muscles are smaller and confined by other muscles, chiefly those operating the legs. Running along the midline of this muscle layer is the heart. If you accidentally remove the muscle layer, the heart will be destroyed, so be careful.
Hemal System
The hemal system is the first organ system uncovered in dorsal dissection (Fig 19). It consists of the hemocoel, blood, and heart. The hemocoel is divided by perforated horizontal membranes, known as diaphragms, into a shallow dorsal pericardial sinus, a spacious perivisceral sinus, and a small ventral perineural sinus (Fig 16-7). Together the sinuses form a large partitioned blood space, the hemocoel. The hemocoel is the functional body cavity of these acoelomate animals.
The heart, surrounded by the pericardial sinus (= pericardial hemocoel) lies immediately under the tergites. Removal of the tergites opens the shallow pericardial sinus, although it is not at all obvious that you are looking at a blood space. It is the space between the exoskeleton and the dorsal diaphragm. The perforated, horizontal dorsal diaphragm is the sheet of connective tissue with longitudinal tergal muscles on top of it (Fig -16-7). This diaphragm separates the pericardial sinus from the much larger perivisceral sinus. These two sinuses are part of the hemocoel. Blood on its way back to the heart flows through the perforations in the diaphragm.
The heart is a longitudinal middorsal tube extending the length of the body in the pericardial sinus and resting on the dorsal diaphragm (Fig 19). The heart has segmental swellings, paired segmental ostia, paired segmental arteries (unusual in insects), and paired segmental alary muscles. The ostia, segmental vessels, and muscles will not be seen and most of the swellings are inconspicuous. The heart opens anteriorly into the cephalic hemocoel of the head and posteriorly bifurcates to form two arteries.
In arthropods blood is pumped anteriorly, during systole, by contractions of circular muscles in the heart wall. Blood leaves the heart, enters the cephalic hemocoel and passes posteriorly through the three sinuses of the hemocoel, eventually passing through the perforations of the diaphragms to enter the pericardial sinus. During diastole contractions of radiating alary muscles cause the heart to dilate and draw blood into its lumen through the ostia. The heart is now refilled with blood and ready to enter another systole. Heartbeat reversal has been observed in cockroaches.
Respiratory System
The respiratory system consists of 10 pairs of spiracles which open into a complex system of distributory tracheae which deliver oxygen to the tissues. In insects the blood is not involved in oxygen transport.
Removal of the tergites exposes the network of branching, tubular, white or silvery tracheae (Fig 19). Use fine forceps to remove muscles and connective tissue covering some of the tracheae and spiracles to improve your view of them.
The tracheae arise at spiracles (Fig 12, 19) in the pleura near the posterolateral corners of the tergites and join with other tracheae to form a single network of air tubes for the delivery of oxygen to the tissues (Fig 21-10B, C). Two pairs of spiracles are present in the thorax and eight pairs in the abdomen. No spiracles are present in the head. The spiracles were seen earlier in your study of the external anatomy.
On each side the respiratory system includes three longitudinal trunks and segmental tracheae to the three hemocoel sinuses and their viscera. The portion of the system you see at present consists, on each side, of a lateral longitudinal trunk extending from spiracle to spiracle (Fig 19), a series of dorsal segmental tracheae extending from the spiracles to the heart and pericardial sinus, and a dorsal longitudinal trunk connecting the dorsal segmental tracheae medially. The dorsal longitudinal trunks lie beside the heart. Other segmental vessels, which cannot be seen at present.
> 1a. Remove a short piece of trachea and make a wetmount with it. Examine it with the compound microscope at 100X and 400X with the light carefully adjusted. Observe the closely spaced rings of chitin, similar in function to the cartilaginous rings in a vertebrate trachea. These rings, known as taenidia, hold the tracheae open, permitting air to pass unimpeded (Fig 21-10A). The taenidia is actually a single helix of chitin, not a series of rings at all. The helix has a very short wavelength so its coils are adjacent to each other, like a Slinky™ at rest. <
As you remove tracheae from the thorax some taenidia may uncoil and extend as a long, tough, very fine thread, thus demonstrating that it is a continuous coil, not a series of separate rings.
Digestive System
" Remove the dorsal diaphragm along with the tergal muscles, tracheae, and heart from the abdomen. Accomplish this by cutting with fine scissors around the periphery of the abdomen. Be very careful that you do not cut deeper than the diaphragm. The crop of the digestive system is a large, thin walled chamber filling most of the abdomen. Its walls may be closely appressed to the dorsal diaphragm, creating the risk that it will be removed with the diaphragm. Remove the muscles, heart, and tracheae, but nothing else, from the thorax. Be careful removing tissue in the thorax that you do not inadvertently destroy the salivary glands and salivary reservoirs, both of which are closely associated with the walls of the anterior gut (esophagus).
The space you have uncovered is the large, spacious perivisceral hemocoel, in which most of the viscera are found (Fig 16-7). Note the large, amorphous, white fat body in the hemocoel (Fig 22). This versatile tissue occupies much of the space in the abdominal hemocoel but is also present in the thorax. It shares many functions with the vertebrate liver and annelid chlorogogen. The fat body functions in storage of lipid, glycogen and protein reserves. Stored food in the fat body supports survival over long periods of starvation and its size varies depending on the extent of starvation. It is a site for intermediary metabolism, amino acid synthesis, blood glucose regulation, vitamin synthesis, and uric acid storage. Some of these functions are mediated by mutualistic, intracellular bacteria known as bacterioids.
Note the two pairs of large diameter tracheae extending through the thorax into the head. One pair lies beside the heart, the other is ventral, beside the esophagus. They will be destroyed as you remove tissues to reveal the gut.
The cockroach gut, like that of other arthropods, consists of an anterior, ectodermal stomodeum, or foregut, a middle endodermal midgut, and a posterior endodermal proctodeum, or hindgut. Both stomodeum and proctodeum, being ectodermal, are lined by epidermis and exoskeleton, which in some regions may be sclerotized. The foregut has chief responsibility for mechanical digestion, trituration, chemical digestion, and storage, the midgut is the region of enzyme secretion, chemical digestion and absorption. Water reclamation, feces formation and storage occur in the hindgut. It is also a storage kidney that sequesters uric acid.
The foregut consists of mouth, pharynx, esophagus, crop, and proventriculus (Fig 21-8A, 16-9). The midgut consists of midgut and digestive ceca. The hindgut is composed of an ileum, colon, and rectum. The ileum and colon together are sometimes known as the intestine.
The mouth was seen earlier opening from the roof of the preoral cavity (Fig 21-7). It opens onto the pharynx which extends dorsally through the head capsule. It will not be seen from your present viewpoint. It becomes the esophagus and exits the head capsule to enter the thorax, where it widens. The esophagus is a conspicuous feature of the anterior thorax (Fig 20) where it can be recognized as a ridged, thin-walled tube.
A pair of white, multi-lobed salivary glands and a pair of bladder-like salivary reservoirs adhere to the walls of the esophagus (Fig 20). Saliva is secreted by the glands and stored in the reservoirs. The salivary glands extend posteriorly from mid- prothorax through the mesothorax. Salivary glands and reservoirs are drained by ducts which extend anteriorly into the head, eventually to coalesce and empty into the salivarium of the preoral cavity. The walls of the ducts are reinforced by helical rings similar to the taenidia of tracheae. Here the saliva is mixed with food particles masticated by the mandible and maxillary lacinia. The saliva contains amylase.
Figure 20. Dorsal view of the perivisceral coelom of a male P. americana. The dorsal diaphragm, heart, and tergal muscles have been removed. Abdominal segments are numbered. Malpighian tubules have been shortened and reduced in number for clarity. Blatt56L.gif
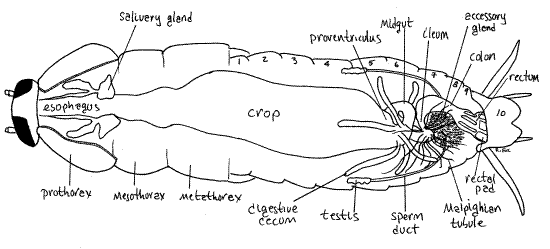
Near the junction of the mesothorax and metathorax the esophagus widens to become the crop. This thin-walled, expansible storage organ may fill the perivisceral hemocoel of the posterior thorax and anterior abdomen. Its thin, transparent walls may push against the body wall and dorsal diaphragm. The crop of preserved specimens may contain a large air bubble. Almost all hydrolysis occurs in the crop making use of enzymes from two sources. Salivary amylase, is mixed with the food in the salivarium and then ingested, whereas enzymes from the digestive ceca, move anteriorly from the midgut.
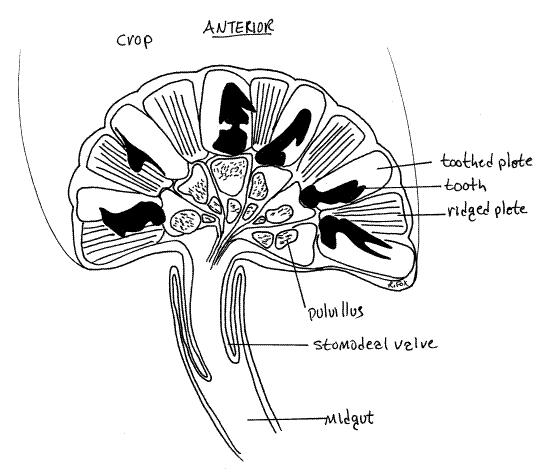
In the vicinity of segments 5-6 the crop narrows to become the short, muscular, thick-walled proventriculus (= gizzard). This region of the foregut has cuticularized walls with heavily sclerotized, black or golden brown teeth (Fig 21, 21-8B). These may be faintly visible through the walls of the crop but the walls of the proventriculus are opaque and you will get a better look later when you open the gut. Esophagus, crop, and proventriculus are regions of the foregut.
The gut narrows abruptly posterior to the proventriculus and becomes the midgut (= ventriculus). The transition from foregut to midgut is marked internally by the stomodeal valve, that protrudes into the anterior midgut (Fig 21).
A whorl of eight long, fingerlike diverticula, the digestive ceca, marks the beginning of the midgut (Fig 20, 21-8A). The ceca increase the surface area for secretion of enzymes and absorption of monomers. Enzymes produced in the ceca include invertase, lipase, maltase, protease, (and perhaps cellulase in some species). Lactase is also present in the midgut. Some of these may be produced by symbiotic gut microorganisms and it may be that amylase is the only digestive enzyme produced by the roach itself.
The midgut makes an asymmetric loop, curving to the right, then bending sharply anteriorly, and then ventrally before heading posteriorly and dorsally. Immediately after resuming its position on the midline it becomes the ileum of the hindgut.
The transition from midgut to hindgut is marked by 60-150 long, slender, white, threadlike, blind-ending Malpighian tubules in six clusters (Fig 21-8A, 21-9). The tubules are hollow gut diverticula that extend into the hemocoel where they are bathed in blood. In Periplaneta they contain intracellular enzymes and may be more important in digestion than in excretion.
The first region of the hindgut is the short, narrow, muscular ileum. It extends for a short distance posteriorly from the midgut and then dilates to become the colon. The transition is marked internally by the ileocolic valve, a sphincter muscle. The colon extends posteriorly to about segment 7 where it expands to become the rectum. The wall of the rectum bears six longitudinal, opaque, white ridges, the rectal pads. The pads protrude into the lumen of the rectum where they remove water from the forming feces. The rectum efficiently reclaims water from feces when water is scarce and conservation is advantageous, producing dry hard fecal pellets. When water is abundant, however, the rectum does not remove water so effectively and produces wet fecal pellets. Insect fecal pellets have characteristic patterns of longitudinal grooves and ridges impressed on them by the rectal pads.
The rectum ends at the anus, under tergite 10 between the two paraprocts. Slip a needle under tergite 10 between the paraprocts and watch it appear in the rectum.
" Use fine scissors to open the posterior crop and proventriculus. Inside the proventriculus you will find an elaborate gastric mill composed of an assortment of sclerotized plates (Fig 21, 21-8A,B). Three sets of six plates each are embedded in the proventricular wall. In a circle adjacent to the crop are 12 plates in two alternating sets. Six are toothed plates, each with a large complex tooth, and six are ridged plates, each with several parallel ridges. The teeth continue the mechanical breakdown of food particles initiated by the mandibles and maxillae. Six additional plates are arranged in a second whorl, this one closer to the midgut. These lightly sclerotized plates support soft cushions, the pulvilli, bearing short fine setae which presumably act as a filter to exclude large particles from the midgut. The astonishing gastric mill of the insect proventriculus is a sight you don’t want to miss.
Continue the incision along the midgut and hindgut noting that these regions are lined by a loose transparent peritrophic membrane (Fig 21-9). If food is present in the gut of your specimen, it will be contained within the peritrophic membrane. The membrane is secreted continuously by the gut epithelium.
Figure 21. Interior of the proventriculus. Blatt57L.gif
Excretory System
Cockroaches may employ several mechanisms for eliminating or inactivating nitrogenous waste products, chiefly as uric acid. It is not known which is most important.
Some nitrogen may be transferred from the blood to the gut lumen, and thus to the feces, by Malpighian tubules and the midgut. In most insects the Malpighian tubules remove wastes and inorganic ions from the blood and secrete them into the gut lumen, where they become incorporated in the feces and are eliminated. Uric acid has not been demonstrated in cockroach Malpighian tubules, however, and it may be that, since they are known to contain digestive enzymes, their role is in digestion rather than excretion.
One of the functions of the fat body is the sequestration, storage, and possibly recycling of nitrogenous waste products. Specialized urate cells in the fat body synthesize uric acid from waste nitrogen and store it indefinitely. Cockroaches on high protein diets develop a hypertrophied fat body functioning as a storage kidney. It is thought that endosymbiotic prokaryotic cells (bacterioids) use the stored nitrogen for protein synthesis. Storage in the fat body is probably the most important excretory mechanism in cockroaches.
In males of a few cockroach species nitrogen is excreted by the accessory glands of the reproductive system. Some tubules of this gland absorb and store uric acid which is then deposited in the spermatophore and eliminated during copulation. This mechanism, of course, is of no use to females and is limited to males of only a few species (such as Blatella germanicabut not Periplaneta).
The periodic molting by arthropods is potentially a mechanism for eliminating unwanted materials from the body. It has been suggested that the nitrogen, as protein, incorporated in the developing cuticle during can serve as a mechanism for eliminating excess nitrogen. The incorporated nitrogen is then jettisoned with the next molt. This mechanism could be effective in nymphs but not adults, since they do not molt.
Reproductive System
Female
The female internal genitalia consist chiefly of a pair of ovaries, two colleterial glands (= female accessory glands) with separate ducts, seminal receptacle (two in some species), and oviducts (Fig 21-11A). Each ovary is a bundle of tapering tubes, the ovarioles, which empty into an oviduct. Oogenesis and gamete maturation occur in the ovarioles. The lateral oviducts, one from each of the two ovaries, join to form a common oviduct that opens through sternite 8 into the genital pouch (Fig 15). The duct from the seminal receptacle opens at the end of the receptacle papilla in the pouch (Fig 23). During copulation the male attaches a spermatophore a sternite in the female genital pouch. Sperm exit the spermatophore, travel up the receptacle duct to the seminal receptacle where they are stored, potentially for a year or more.
" The female reproductive system will not be visible until much of the fat body in the dorsal abdomen has been removed. Begin first in the posterior abdomen and remove tergites 7-9 if you have not already done so. The fat body covers and is packed around the colleterial glands and ovaries and must be removed carefully without damaging either. The size of the fat body varies depending on metabolic and reproductive condition of the insect. Reserves stored in the fat body are transferred to the developing eggs when the female is reproductive active.
Once you have removed the fat body, the left colleterial gland will be conspicuous as spaghetti-like tangle of many long, slender, branching, opaque white tubes. The glands lie beside and dorsal to the hindgut and, along with the fat body, obscure your view of everything else in the posterior abdomen (Fig 22). The colleterial gland is not restricted to the surface and fills much of the space between the sternum and tergum. The smaller right colleterial gland will probably not be seen.
The two colleterial glands empty via separate ducts into the genital pouch. The two glands differ dramatically both morphologically and chemically. The left gland is much larger than the right and completely obscures it from view. Secretions from both are released simultaneously into the genital pouch and react to form the sclerotized lining of the pouch that will become the ootheca. Protein released from the left gland is tanned by phenol from the right gland. The resulting wall of the ootheca wall is composed of scleroprotein identical to that in cuticular sclerites, although chitin is absent. Eggs are deposited into this lining as they leave the ovarioles and the lining then hardens (tans) to become the ootheca.
The ovaries lie laterally in the abdomen, one on either side of the gut and may extend for most of the length of the abdomen. Or, if immature or inactive, may be restricted to abdominal segments 5-7. The ovaries are hidden from view by the fat body and colleterial glands, which much be removed.
Each ovary is a bundle of large tapering tubes, the ovarioles. In Periplaneta each ovary consists of eight ovarioles (Fig 22). Each ovariole is a connective tissue tube consisting of two regions. The germarium is the short, small-diameter, upstream, anterior region. Oogonia in the germarium initiate oogenesis culminating downstream in the production of oocytes. Thevitellarium is the much longer, much wider, downstream region in which follicle cells transfer nutriment to the maturing and growing oocytes. The diameter of the ovariole is very small in the germarium but increases downstream as the developing gametes accumulate yolk from the follicle cells and increase dramatically in size (Fig 22).
Figure 22. Dorsal view of the opened abdomen of a female P. fuliginosa. The fat body and colleterial gland have been removed from the left side to reveal the left ovary. T = tergite. Blatt58L.gif
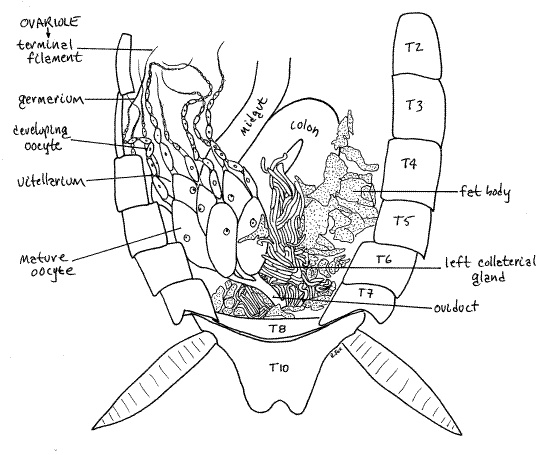
The downstream ends of the eight ovarioles of each ovary coalesce to form a lateral oviduct (Fig 21-11A). Remove the colleterial glands from one side and find the lateral oviductexiting the large, posterior, basal end of the ovary. Upon exiting the ovary the oviduct passes deep into the tissues in the ventral abdomen and extends medially to join the other oviduct on the ventral midline. The junction of the right and left lateral oviducts forms the short unpaired common oviduct which passes posteriorly to open through the female gonopore in the genital pouch (Fig 15, 23).
Maturing gametes are lined up one after another in the ovariole with oocytes maturing as they move downstream (Fig 22). The mature egg (actually, it is a secondary oocyte that will not complete meiosis until it is fertilized) is the downstream-most gamete in the ovariole. Each ovariole typically has a single mature egg ready for fertilization and incorporation into an ootheca (Fig 22). Eggs leave the ovarioles via the oviduct and are fertilized by sperm from the seminal receptacle as they enter the genital pouch. The ovipositor manipulates them and orients them properly in the developing ootheca. If the eggs are mis-oriented, the nymphs will be unable to escape from the ootheca and will die . The ootheca hardens around two rows of eight eggs each, one from each of the 16 ovarioles (eight ovarioles in each of two ovaries). The number of eggs in the ootheca depends on the number of ovarioles in the ovary and differs with species.
Females of some species (e.g. Blatella germanica) retain the ootheca and brood their eggs. Most, including Periplaneta, drop the completed ootheca and abandon it.
Figure 23. Dorsal view of the floor of the genital pouch with the gut, fat body, and colleterial glands removed. Blatt59L.gif
Male
The male reproductive system consists of paired testes, each draining by a sperm duct (= vas deferens) to a common ejaculatory duct which opens via the male gonopore in the genital pouch (Fig 21-12A). Associated with the confluence of the two sperm ducts are two clusters of secretory accessory glands. Near the confluence, the sperm ducts are expanded to form seminal vesicles. It is thought that spermatogenesis occurs in the testes during the last nymphal instar and the spermatozoa stored for later use as an imago.
The testes are located dorsolaterally in segments 4 and 5 (Fig 20). They are embedded in the white fat body and can be difficult or impossible to demonstrate in adults, especially older specimens. Part of this difficulty is attributed to the alleged (disputed) degeneration of the testes in older adults but most of the problem is due to the fat bodies. Like the testes, the sperm ducts are difficult to demonstrate and you probably will not see them.
Prior to copulation a spermatophore is formed in the male ejaculatory duct when secretions of the accessory glands enclose and harden around a mass of sperm from the seminal vesicles. During copulation the phallomeres open the female’s genital pouch and hold the male’s genitalia in the correct position. The ejaculatory duct on the ventral phallomere is held adjacent to the opening of the spermathecal papilla (Fig 23) and the spermatophore released. It is believed that the spermatophore is glued in place by secretions of the male’s phallic gland. In about 24 hours or less, sperm exit the spermatophore and are stored in the female’s seminal receptacle.
Figure 24. Dorsal dissection of the posterior hemocoel and genital pouch of a male P. americana. The internal genitalia (accessory glands) have been removed. Blatt60L.gif
" In dorsal dissection your view of the male reproductive system is probably obscured by lobes of the bright white fat body. Remove this region of the fat body and tergite 8-9 if you have not already done so.
Following removal of the fat body, the male accessory gland (= mushroom gland) is by far the most conspicuous of the male's internal genitalia (Fig 20). It secretes the covering of the spermatophore and is a mass of worm-like diverticula in the posterior hemocoel under tergites 7-8 beside the anterior end of the posterior colon and anterior rectum. The mass includes large (utriculi majores) and small (utriculi breviores) diverticula and is usually surrounded by lobes of the fat body.
The two sperm ducts enter the gland and the seminal vesicles are enclosed in the mass of tubules. The thick ejaculatory duct, formed by the union of the sperm ducts, exits ventrally from the center mass of accessory glands and extends deep into the ventral body wall and then posteriorly to the gonopore on the dorsal surface of the ventral phallomere (Fig 24).
Most of the phallic gland is ventral to the nerve cord and should not be exposed until you have completed your study of the nervous system (Fig 24). It is deeply ventral in the posterior abdominal cavity, ventral even to the ventral diaphragm and is a little to the right of the midline. Remove the tissue ventral to the posterior nerve cord to expose the gland. Its appearance is distinctive. It is glandular, white, and shaped like an elongate teardrop, tapering posteriorly to form the phallic gland duct. It is between abdominal ganglion 5 and the terminal ganglion. It is ventral to the terminal ganglion and the right cercal nerve. Its duct opens in the genital pouch between the pseudopenis and acutolobus of the left phallomere (Fig 18, 24). During copulation secretions of the phallic gland are used to attach the spermatophore in the vicinity of the opening of the seminal receptacle.
Nervous System
The basic features of the insect nervous system are the dorsal brain joined by circumenteric connectives to ventral nerve cord with paired segmental ganglia. It includes both somatic and visceral components and is served by a variety of sensory devices. Most, but not all, segments have a pair of segmental ganglia. Segmental ganglia and the brain consist of paired lateral ganglia connected by a transverse commissure. In cockroaches the commissures are short resulting in fusion of the lateral ganglia so each segmental ganglion seems to be a single ganglion.
" Remove the gut, ventral diaphragm, and muscles from the thorax and abdomen. Remove the longitudinal body wall muscles (sternal muscles) and connective tissue as necessary from the floor of the abdominal cavity to reveal the ventral nerve cord (Fig 16-11). The cord consists to two side-by side, parallel, longitudinal connectives which are united by transverse commissures passing between paired segmental ganglia. Notice the longitudinal ventral tracheal trunk lying beside the ventral nerve cord.
Three thoracic ganglia are located in the thorax (Fig 25). From each extends several pairs of nerves to the abundant muscles of these segments. These are large ganglia.
Figure 25. Dorsal dissection of P. americana. T = thoracic ganglion, A = abdominal ganglion. The abdominal segments are numbered. Blatt61L.gif
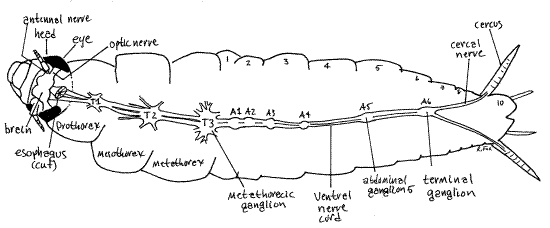
In the abdomen are six much smaller abdominal ganglia. Abdominal ganglia 1-3 are scarcely wider than the double nerve cord and can be difficult to distinguish from the cord. Abdominal ganglia 4 and 5 are usually readily discernable. The sixth abdominal ganglion, known as the terminal ganglion, is the largest of the abdominal ganglia and is easily recognized. The nerve cord ends at the terminal ganglion which gives rise to two large sensory cercal nerves to the cerci. The terminal ganglion is the coalesced ganglia of several posterior segmental ganglia.
" Using your finest scissors and forceps carefully remove the epicranium from the region between the compound eyes and antennae. Remove muscles as necessary to reveal the bright white, dorsal brain between the compound eyes (Fig 25). The brain consists of two cerebral ganglia fused across the midline. Short thick optic nerves extend from the eyes to the brain. Smaller antennal nerves run from the antennae to the brain. Remove the mandibles and other tissues from one side of the head to reveal the thick circumesophageal connectives.These exit the ventral side of the brain and extend posteriorly to pass around the esophagus, and then coalesce to form the subesophageal ganglion ventral to the gut. Nerves to the mandibles, maxillae, and labium exit the subesophageal ganglion. A pair of longitudinal nerve cords exit the subesophageal ganglion as the ventral nerve cord and pass posteriorly through the foramen magnum to thoracic ganglion 1 (Fig 25).
References
Anon. 1967. Pictorial keys, arthropods, reptiles, birds, and mammals of public health significance. US Public Health Service, Communicable Disease Center, Atlanta. 192 pp.
Barbara KA . University of Florida Institute of Food and Agricultural Sciences, Periplaneta americana page: http://creatures.ifas.ufl.edu/urban/roaches/american_cockroach.htm
Bell WJ, Adiyodi KG. 1981. The American cockroach. Chapman and Hall, London.
Borror DJ, Triplehorn CA, Johnson NF. 1986. An introduction to the study of insects. Saunders, Philadelphia. 875pp.
Buck JB, Keister ML. 1950. Periplaneta americana, pp475-496 in Brown FA (ed), Selected invertebrate types. Wiley, New York. 596pp.
Cornwell PB. 1968. The cockroach, vol 1, A laboratory insect and industrial pest. Hutchinson of London. 391pp.
Ebling W. 1975. Urban entomology. Univ. Calif, Richmond CA.
Furman DP, Catta EP. 1970. Manual of medical entomology, 3 rd ed. National Press Books, Palo Alto. 163 pp.
Guthrie DM, Tindall AR. 1968. The biology of the cockroach. Edward Arnold Publisher. 408pp.
McKittrick FA. 1964. Evolutionary studies of cockroaches. Cornell Univ. Agric. Expt. Sta. Mem. 389.
Miall LC, Denny A. 1886. The structure and life history of the cockroach. Lovell Reed Company, London. 224pp.
Ruppert EE, Fox RS, Barnes RB. 2004. Invertebrate Zoology, A functional evolutionary approach, 7 th ed. Brooks Cole Thomson, Belmont CA. 963 pp.
Snodgrass RE. 1935. Principles of insect morphology. McGraw-Hill, New York. 667pp.
Stang LA. University of Florida Institute of Food and Agricultural Sciences, Evania appendigaster page:
http://creatures.ifas.ufl.edu/urban/roaches/ensign_wasp.htm
Supplies
Dissecting microscope
Living or preserved adult cockroach, preferably Periplaneta
Dissecting equipment with microdissecting forceps
Small dissecting pan made from anchovy or sardine can with dissecting wax bottom.
Anesthetic (chloroform, ether, or carbon dioxide) or
Killing agent (ethyl acetate)
# 1 stainless steel insect pins