Invertebrate Anatomy OnLine
Obelia ©
Hydrozoan
30jun2006
Copyright 2001 by
Richard Fox
Lander University
Preface
This is one of many exercises available from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises can be accessed by clicking on the links on the left. A glossary and chapters on supplies and laboratory techniques are also available. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Cnidaria P, Medusozoa, Hydrozoa C, Leptothecatae O, Campanulariidae F (Fig 7-75, 7-74)
Cnidaria P
The cnidarian body consists of a central blind sac, the coelenteron (= gastrovascular cavity), enclosed by a body wall comprising two epithelia, the outer epidermis and the inner gastrodermis (Fig 7-1, 7-2). A gelatinous connective tissue layer, the mesoglea, lies between the two epithelia. The mouth opens at one end of the coelenteron and marks the oral end. The mouth is at the tip of a process, the manubrium that elevates it above the oral surface. The opposite pole is the aboral end. The imaginary line connecting the oral and aboral poles is the axis of symmetry around which the radial symmetry of the body is organized. The mouth is usually surrounded by one or more circles of tentacles.
The defining cnidarian feature is, of course, their possession of stinging cells, or cnidocytes (Fig 7-8). Characteristic of the epidermis, they are also sometimes found in the gastrodermis. Cnidocytes contain an explosive organelle, the cnida, which, upon proper stimulation, inverts and ejects a slender, often barbed and toxic thread in the direction of prey or predator (Fig 7-9). Three types of cnidae are found in cnidarians (Fig 7-10). Nematocysts (in nematocytes), spirocysts (in spirocytes), and ptychocysts (in ptychocytes). All toxic cnidae are nematocysts whereas spirocysts are sticky, and the everted tubules of ptychocysts are used for constructing feltlike tubes. Most cnidae are nematocysts and these are present in all three higher cnidarian taxa. Spirocysts and ptychocysts are found only in Anthozoa.
The basic body plan described above can be manifest as a swimming medusa or attached polyp. In some taxa only one generation is present whereas in others both are found. A life cycle featuring alternation of sexual, swimming medusae with benthic asexual polyps is typical of many cnidarians.
All cnidarians are carnivores feeding on live prey which they usually capture using tentacles armed with cnidocytes. Digestion occurs in the coelenteron which is typically equipped with ciliated canals for distribution of partly digested food. Cnidarians are ammonotelic and diffusion across the body and tentacle surface eliminated the ammonia from the body. Gas exchange is across the general body surface. The nervous system is a plexus of basiepithelial neurons serving sensory and motor systems (Fig 7-6). Most cnidarians are gonochoric. The life cycle typically includes a planula larva. Cnidarians are chiefly marine but the well-known Hydra is an exception.
Medusozoa
Medusozoa comprises those cnidarians whose life cycle includes a medusa generation that alternates with a polyp generation (Fig 7-75B). Symmetry is radial and tetramerous. Nematocysts are the only type of cnidocyte present. Included taxa are Scyphozoa (jellyfishes) and Hydrozoa (hydroids, Hydra, Portuguese men of war, etc).
Hydrozoa C
Hydrozoa is a diverse taxon of about 3000 species of mostly marine cnidarians. The life cycle usually includes both polyp and medusa generations (Fig 7-65A) but may be entirely polyp (Fig 7-65B) or entirely medusa (Fig 7-65C). Polyps typically are colonial and medusae usually solitary. Some form colonies of combinations of polyps and medusae. The few freshwater cnidarians, such as Hydra, Vallentinia, and Craspedacusta, are hydrozoans.
Hydrozoan polyps are usually small, about 1 mm in length, and colonial. Hydromedusae are also small, at least in comparison with scyphomedusae, and are usually less than 1 cm in diameter. Hydromedusae are further distinguished from scyphomedusae by possession of a velum, a circumferential shelf of tissue that encircles the subumbrellar concavity and functions as an adjustable diaphragm to create a pulse of water for swimming. Medusae are tetramerously symmetrical as are scyphomedusae and four radial canals and four tentacles are usually present.
Cnidocytes are found only in the epidermis and germ cells arise in the gastrodermis but may, or may not, migrate to the epidermis to form gonads and mature into gametes. Gonads may be on the manubrium or on the radial canals. Gametes, even if gastrodermal, are released directly to the surrounding water, never, in contrast with scyphomedusae, into the coelenteron.
Leptothecatae O
Both polyps and medusa generations in the life cycle (Fig 7-63). Medusae are thick cup-shaped and large for hydromedusae. The polyps are colonial, thecate, and tiny but the medusae, which are low and flat, are sometimes large, up to 50 cm. Statocysts, but not ocelli, are present and the gonads are on the radial canals.
Laboratory Exercise
1. Obelia is representative of typical hydrozoan morphology and possesses a life cycle in which polyp and medusa are of approximately equal importance. There are several species ofObelia and any can be used for this exercise. If living specimens are available they should be studied in seawater. Most laboratories, however, will provide commercially prepared wholemount slides.
Polypoid Generation
Place a commercially prepared slide on the stage of the compound microscope and focus at 40X.
Figure 1. Obelia polyp generation. A small portion of a colony. Hydrozoa4La.gif
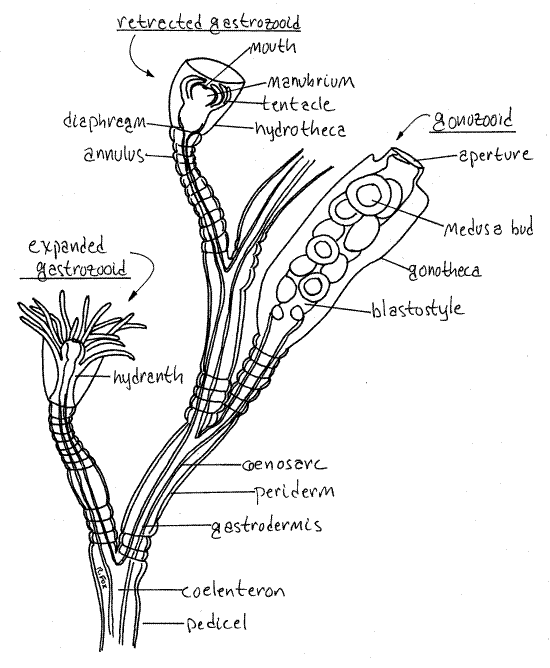
Obelia colonies consist of numerous polyps on branched stalks (Fig 1, 7-64A). A creeping, rootlike stolon lies on the surface of a firm substratum and serves as anchor and base for the entire colony. Commercially prepared slides include only a small piece of the colony and the stolon is rarely present. It may be absent from living or preserved material also, unless care has been taken to collect it. Numerous erect pedicels (= stem) arise from the stolon. The stems branch in patterns characteristic of the species and bear polyps.
Obelia polyps (= zooids) are dimorphic, there being two types in the colony. Gastrozooids are responsible for feeding, which they accomplish by capturing and ingesting zooplankton. Gonozooids are reproductive and produce medusae by asexual budding. Gastrozooids look like little flowers and have a circle of stinging tentacles surrounding a mouth, which opens into the coelenteron. Gonozooids are elongate, lack tentacles or mouth, do not feed, and do not look like flowers.
The stems, stolons, and polyps (Fig 1) are enclosed in a transparent, chitinous exoskeleton called the periderm (= perisarc). This is a non-living secretion of the epidermis. Inside is living tissue, collectively known as the coenosarc (pronounced SIN o sark, coen = common or together, sarc = flesh) consisting of the outer, nonciliated epidermis and the inner, ciliatedgastrodermis with the thin, acellular mesoglea in between. The three layers can be distinguished by careful focusing with 400X. The epidermis and gastrodermis are relatively thick and the mesoglea is a narrow dark line between them.
The hollow interior of the animal is the coelenteron, or gastrovascular cavity. The cavity extends uninterrupted throughout the colony and is both a chamber for extracellular digestion and a fluid transport system to deliver partially digested food through the colony.
Gastrozooids
The gastrozooids provide food for the entire colony, including the nonfeeding pedicels, stolons, and gonozooids. Find a gastrozooid and study it carefully. The periderm of its stem is annulated, or ringed, each ring being an annulus. The polyp is enclosed in a bell-shaped expansion of the perisarc called the hydrotheca (theca = a case). Polyps that are enclosed in this manner are said to be thecate. Obelia, being thecate belongs to the taxon Leptothecatae. Some hydrozoan species (Anthoathecatae) have athecate polyps.
The coenosarc of the gastrozooid is called a hydranth (= water flower). It rests on a transverse shelf of periderm, the diaphragm, at the base of the hydrotheca. The coenosarc of the pedicel passes through a small hole in the center of the diaphragm and is continuous with that of the hydranth. The distal, or free, end of the hydranth bears a single circle of filiform tentaclesused to capture prey, sting it, subdue it, and transfer it to the mouth. The tentacles are solid, contractile, and armed with cnidocytes. Cnidocytes are clustered together in batteries that appear as small bumps on the tentacles.
In the center of the whorl of tentacles is the erect manubrium, which bears the mouth at its center. The mouth opens into an expanded region of the coelenteron known as the stomach. The stomach connects with the canal-like coelenteron that extends throughout the colony. The gastrodermis of the stomach contains secretory cells that release proteases. These enzymes partially hydrolyze food into small particles and molecules for distribution by ciliary currents to other parts of the colony. Particles are phagocytized by cells lining the coelenteron and digestion is completed intracellularly.
Gonozooids
The gonozooids are reproductive polyps. They may be absent from some colonies. They arise from the pedicel at the bases of the gastrozooids and consist of an elongate cylinder of perisarc, called the gonotheca, which, like the hydrotheca, is open distally to the sea. The gonotheca encloses a cylinder of coenosarc called the blastostyle. The blastostyle is the living part of the gonozooid. In mature individuals its surface bears small medusa buds which are produced asexually. These eventually develop into medusae and break free from the blastostyle to swim out the aperture of the gonotheca into the sea.
Medusoid Generation
Hydrozoan medusae, known as hydromedusae, are small, saucer-shaped, free-swimming, sexually reproducing individuals.
Figure 2. Obelia medusa. Hydrozoa5L.gif
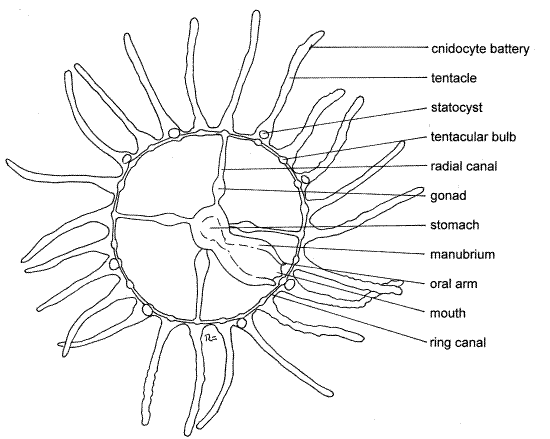
Place a commercially prepared slide of Obelia medusae on the stage of the compound microscope and focus at 40X. The medusae of prepared slides are stained a variety of meaningless colors but in life they are transparent and colorless.
Look at the slide without the benefit of the microscope and note the size of the medusae. Hydromedusae are, in general, much smaller than scyphomedusae and those of Obelia are even smaller than usual.
The saucer-shaped Obelia medusae are flatter than most hydromedusae but those of commercial preparations are further flattened by the weight of the coverslip (Fig 2). Most of the specimens on your slide will probably be lying flat on either the oral or aboral surface. A few specimens may be folded and in side view.
Hydromedusae have a distinct tetramerous (4-part) radial symmetry with most structures occurring in fours or multiples of four.
The arched saucerlike body is referred to as the bell. It has a convex upper, exumbrella (aboral surface ) and a concave lower, subumbrella (oral surface). The aboral surface is smooth and rounded. In Obelia, the velum, which is characteristic of most hydromedusae, is rudimentary and is not visible. Numerous tentacles extend from the margin of the bell. The tentacles contract when preserved and those of living specimens would be much longer.
In life, the manubrium hangs like a pendulum into the center of the subumbrellar space below the oral surface but in wholemounts it is deflected to the side (Fig 2). It may be necessary to look at several medusae to get a good side view of the manubrium.
The mouth is in the center of the free, oral end of the manubrium and is flanked by four short oral arms. The manubrium and oral arms are well supplied with cnidocytes. Take optical sections through the manubrium to convince yourself that it is hollow.
The coelenteron consists of the stomach at the base of the manubrium, where extracellular digestion occurs, and a set of ciliated canals used to distribute partly digested food. The mouth opens into the spacious lumen of the manubrium which in turn empties into the stomach. The stomach is in the center of the bell and usually stains darker than the surrounding tissue.
Four radial canals extend from the stomach to the periphery of the bell where they join the circular ring canal in the margin of the bell. The radial canals are usually faint and indistinct in commercial preparations but are visible with careful focusing with good light adjustment. The mesoglea of the exumbrella is thick and accounts for most of the thickness of the bell. That of the subumbrella is thin. Consequently, the radial canals are easier to see in subumbrellar view.
A distinct, darkly staining, epidermal gonad, either ovary or testis, can be seen below each radial canal near the stomach.
Twenty to thirty solid tentacles radiate from the edge of the bell. The tentacle epidermis has abundant sensory cells and cnidocytes. The tentacles of preserved specimens are contracted and much shorter than those of relaxed living specimens. The tentacles are often broken in commercial preparations. The cnidocytes are concentrated in rings, or batteries, and are used to sting and subdue the prey. A swollen tentacular bulb in the base of each tentacle is the site of intracellular digestion and cnidocyte formation.
There are eight spherical statocysts spaced evenly around the margin of the bell. Each is at the base of a tentacle. The statocysts are hollow, thin-walled, epidermal vesicles containing a calcareous statolith and a ciliated sensory epithelium. There are no ocelli in Obelia or other Leptothecatae.
References
Brown FA (ed.) 1950. Selected Invertebrate Types. Wiley, New York. 597p.
Bullough. WS. 1958. Practical Invertebrate Anatomy (2 nd ed). MacMillan, London. 483p.
Ruppert EE, Fox RS, Barnes RB. 2004. Invertebrate Zoology, A functional evolutionary approach, 7 th ed. Brooks Cole Thomson, Belmont CA. 963 pp.
Thomas MB, Edwards NC. 1991. Cnidaria: Hydrozoa. Pp 91-183 in Harrison FW, Westfall JA (eds). Microscopical Anatomy of Invertebrates, vol. 2. Wiley-Liss, New York.
Supplies
1 compound microscope
1 wholemount of Obelia polyps
1 wholemount of Obelia medusae