Invertebrate Anatomy OnLine
Novobisium carolinense ©
Pseudoscorpion
18jun2006
Copyright 2001 by
Richard Fox
Preface
This is one of many exercises available from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises can be accessed by clicking on the links to the left. A glossary and chapters on supplies and laboratory techniques are also available. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Arthropoda P, Chelicerata sP, Euchelicerata, Arachnida C, Pseudoscorpiones O, Neobisiidae F (Fig 16-15, 18-47)
Arthropoda P
Arthropoda, by far the largest and most diverse animal taxon, includes chelicerates, insects, myriapods, and crustaceans as well as many extinct taxa such as Trilobitomorpha. The segmented body primitively bears a pair of jointed appendages on each segment. The epidermis secretes a complex cuticular exoskeleton which must be molted to permit increase in size. Extant arthropods exhibit regional specialization in the structure and function of segments and appendages but the ancestor probably had similar appendages on all segments. The body is typically divided into a head and trunk, of which the trunk is often further divided into thorax and abdomen.
The gut consists of foregut, midgut, and hindgut and extends the length of the body from anterior mouth to posterior anus. Foregut and hindgut are epidermal invaginations, being derived from the embryonic stomodeum and proctodeum respectively, and are lined by cuticle, as are all epidermal surfaces of arthropods. The midgut is endodermal and is responsible for most enzyme secretion, hydrolysis, and absorption.
The coelom is reduced to small spaces associated with the gonads and kidney. The functional body cavity is a spacious hemocoel divided by a horizontal diaphragm into a dorsal pericardial sinus and a much larger perivisceral sinus. Sometimes there is a small ventral perineural sinus surrounding the ventral nerve cord.
The hemal system includes a dorsal, contractile, tubular, ostiate heart that pumps blood to the hemocoel. Excretory organs vary with taxon and include Malpighian tubules, saccate nephridia, and nephrocytes. Respiratory organs also vary with taxon and include many types of gills, book lungs, and tracheae.
The nervous system consists of a dorsal, anterior brain of two or three pairs of ganglia, circumenteric connectives, and a paired ventral nerve cord with segmental ganglia and segmental peripheral nerves. Various degrees of condensation and cephalization are found in different taxa.
Development is derived with centrolecithal eggs and superficial cleavage. There is frequently a larva although development is direct in many. Juveniles pass through a series of instars separated by molts until reaching the adult size and reproductive condition. At this time molting and growth may cease or continue, depending on taxon.
Chelicerata sP
Chelicerata is a large taxon that includes spiders, scorpions, pseudoscorpions, ticks, mites, horseshoe crabs, sea spiders, and many others. The group originated in marine habitats but almost all modern chelicerates are terrestrial.
The body is divided into an anterior cephalothorax with six pairs of appendages and a posterior abdomen which, in most groups, does not bear appendages or has highly modified appendages. The first appendages of the cephalothorax are the chelicerae. Antennae are not present and the brain has two regions rather than the three found in mandibulates. Appendages are primitively biramous but are uniramous in almost all Recent taxa.
Euchelicerata
The segments of the cephalothorax are fused and covered by a dorsal shield, or carapace. Two median eyes are present.
Arachnida C
Arachnids are the terrestrial descendents of the early aquatic chelicerates. The taxon includes the mites, scorpions, pseudoscorpions, spiders, harvestmen, and several other taxa. All are adapted for a terrestrial existence with internal gas exchange and a waterproof integument. Some are capable of silk production and many use toxins to subdue or kill the prey. Most are carnivores, digestion is usually outside the body, and food is liquefied before ingestion.
Pseudoscorpiones O
Pseudoscorpions are tiny arachnids rarely exceeding 7 mm in length and usually less than half that. They are widespread and common in leaf litter, under bark, and in moss and debris where they are seldom noticed. One species lives in human habitations where it is known as the "book scorpion".
Pseudoscorpions are a distinct and homogenous group with no close ties to other arachnid taxa. Their resemblance to true scorpions is superficial and is due to their common possession of large crab-like pedipalps.
Pseudoscorpions are carnivores and prey on other small arthropods. They move rapidly backwards although some move forwards also. Some move sideways. They exhibit phoresy in which they attach with the pedipalps to the leg of a flying insect and hitch a ride to a new habitat.
Pseudoscorpions have a cephalothorax with a solid undivided carapace and an abdomen with 12 distinct segments. They have relatively small chelicerae and enormous pedipalps. Both are chelate. The chelicerae have silk glands and spin silk whereas the pedipalps have poison glands. There are the usual four pairs of legs on the cephalothorax. The abdomen does not bear appendages. The junction between cephalothorax and abdomen is broad, usually as wide as the cephalothorax although in some species the cephalothorax narrows slightly as it joins the abdomen. There is no narrow pedicel connecting the cephalothorax and abdomen as there is in spiders. Pseudoscorpions are gonochoric and courtship behavior may be elaborate.
Laboratory Specimens
Pseudoscorpions are easily provided for a laboratory exercise by extracting leaf litter with a Berlese funnel (see Techniques chapter). In this technique a handful or two of leaf litter is placed in a large funnel and a 75-watt light bulb left burning for several days at the top of the funnel. A vial of water or alcohol is attached to the small end of the funnel. The heat and desiccation from the lamp drives the animals downward in the funnel until they drop into the vial from which they can be recovered and used in the lab. The technique will yield a variety of small soil arthropods, typically including mites, many insects, millipedes, centipedes, collembolans, pseudoscorpions, and many others.
Because of the homogeneity of the taxon, any species of pseudoscorpion can be used with this anatomical account.
External Anatomy
Place the largest pseudoscorpion you can find in a 6-cm culture dish of water on the stage of the dissecting microscope. Preserved specimens are better examined in water than in alcohol. Convection currents in alcohol can cause these small animals to drift annoyingly about the dish. The pseudoscorpion cuticle, however, resists wetting and some species are nearly impossible to immerse in pure water. Instead they float on the surface tension. This can be corrected by adding a tiny (!) bit of detergent to the dish. Simply touch the tip of the forceps to a bar of soap or to liquid detergent and then touch the surface of the water in the dish. The surfactant makes it possible to sink the specimens, although they retain a tendency to become captured in the surface film.
Examine the specimen with medium to high power of the dissecting microscope. Manipulate it with fine forceps and microneedles. The body, like that of other arachnids, is divided into an anterior cephalothorax and a posterior abdomen (Fig 1, 18-32). The pseudoscorpion cephalothorax is unsegmented whereas the abdomen is distinctly segmented. The cephalothorax joins the abdomen without narrowing significantly and there is no pedicel.
Cephalothorax
Dorsally and laterally the cephalothorax is completely covered by the carapace, a large darkened sclerite of hardened exoskeleton. Laterally the cephalothorax is covered by the overhang of the carapace and the soft, unsclerotized pleural membrane. In general, sclerites, of which the carapace is but one of many, are darker than the softer unsclerotized exoskeleton surrounding them.
Dorsally Novobisium has two pairs of white lateral eyes at the anterolateral corners of the carapace (Fig 1). Other species may have one pair or no eyes.
The cephalothorax bears the usual six pairs of appendages of the typical arachnid. Anteriorly there is a pair of short, chelate chelicerae (Figs 1, 2, 3). These are elaborate structures equipped with silk-spinning spinnerets. The chelicerae consist of only two articles. The proximal article forms the base of the chela and the immovable finger (Fig 1). The smaller distal article articulates with the first distally and is the movable finger. The fingers oppose each other and form a prehensile chela that may be used to masticate the prey or to simply tear a hole in it.
Figure 1. Dorsal view of Novobisium carolinense from leaf litter collected in Greenwood, South Carolina. C = coxa, f = femur, m = metatarsus. ta = tarsus, ti = tibia, tr = trochanter. Pseudo10L.gif
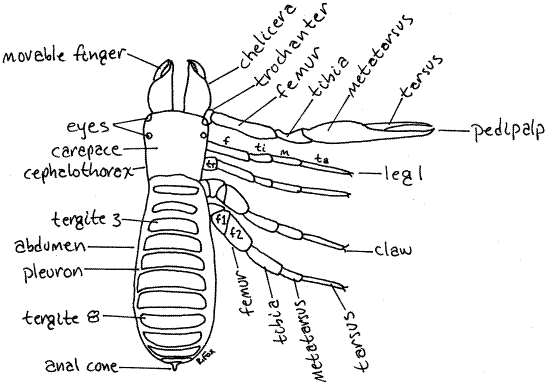
The second pair of appendages is the very large and conspicuous pedipalps (Fig 1). Like the chelicerae, the pedipalps are chelate. They comprise six articles, the last two of which form the chela. The articles of the pedipalp, from proximal to distal, are the coxa, trochanter, femur, tibia, metatarsus, and tarsus. The tarsus is the movable finger of the chela. The pedipalps are used to capture food, nest building, courtship, fighting, and sensory reception.
The remaining four pairs of appendages are walking legs. The walking legs form two groups. Legs 1 and 2 are relatively small and resemble each other whereas 3 and 4 are larger and similar to each other. The legs typically consist of six or seven articles (coxa, trochanter, femur, tibia, metatarsus, and tarsus) but there are many different ways in which these basic articles can be reorganized. In Novobisium (Figs 1, 2, 3) the femurs of legs 3 and 4 are subdivided so that each leg has two femora and a total of 7 articles. Legs 1 and 2 have undivided femora and a total of six articles. Two tiny tarsal claws can be seen at the distal end of each tarsus (Fig 1). In many pseudoscorpions the tarsus is subdivided.
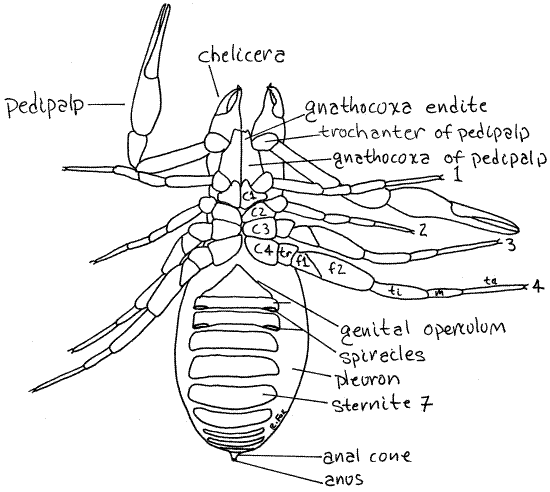
Ventrally the cephalothorax is covered by the five pairs of coxae belonging to the pedipalps and the four pairs of legs (Fig 2). The coxae of the pedipalps are very large and are known as the gnathocoxae. Each gnathocoxa has an anterior process, the endite, that forms the floor of the preoral cavity from which the mouth opens.
The coxae of the walking legs are relatively small. The coxae are not fused with each other but neither are they movable. The external openings of the coxal glands are located medial to the coxae of leg 3 (Fig 2). These are small and difficult to see.
Figure 2. Ventral view of Novobisium. C = coxa, f = femur, m = metatarsus. ta = tarsus, ti = tibia, tr = trochanter. Pseudo11L.gif
Abdomen
The pseudoscorpion abdomen consists of 12 segments. Like scorpions, but unlike other arachnids, the abdomen is conspicuously segmented. Pseudoscorpions have no abdominal appendages.
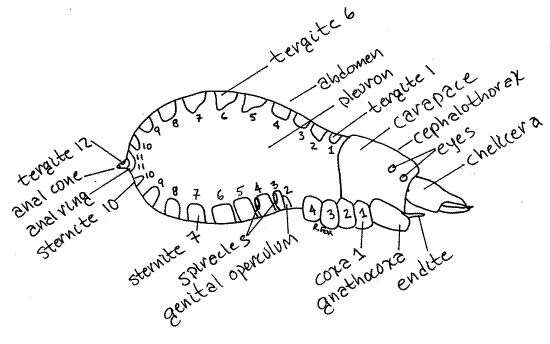
Dorsally the 12 abdominal segments are covered by 11 conspicuous sclerites, known specifically as tergites (Figs 1, 3). Each tergite may be divided into right and left halves or it may be a continuous single piece as in Novobisium (Fig 1). The tergites are surrounded by unsclerotized exoskeleton, which is soft, flexible, and pale.
Ventrally the abdomen bears another set of sclerites, these known as sternites, on segments 2-11. Segment 1 lacks a sternite. The sternites of segments 2 and 3 are fused to form thegenital operculum. The two genital apertures, or gonopores, open through the genital operculum.
Abdominal segment 12 is the tiny anal cone that projects posteriorly between tergite 11 and sternite 11 (Fig 3). It is very small and inconspicuous and can be retracted into the 11 thsegment. The tiny 12 th tergite is on the anal cone. The anus is located at its tip. In taxa tergite 11 and sternite 11 are fused to form a ring around the anal cone.
Two pairs of spiracles connect the tracheal system with the atmosphere and are located laterally on the posterior edges of sternites 3 and 4 (Fig 2).
Figure 3. Lateral view of Novobisium. Coxae, sternites, and tergites are numbered. Pseudo12L.gif
Internal Anatomy
Pseudoscorpions are far too small for a laboratory study of gross internal anatomy.
Digestive System
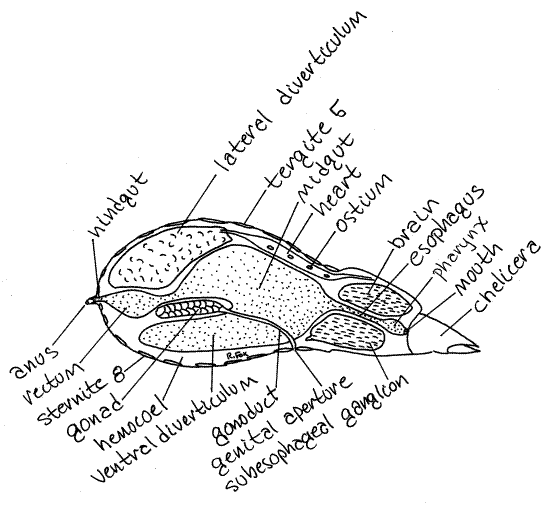
The mouth is on the posterior wall of the preoral cavity located between the bases of the chelicerae and dorsal to the gnathocoxae of the pedipalps. The mouth opens into a short muscular pharynx, or foregut (Fig 4, 18-33). A long esophagus connects the pharynx with the midgut. The esophagus is also part of the foregut. The large and important midgut with several large digestive ceca occupies most of the interior of the abdomen. It is the site of hydrolysis and storage. Glycogen and lipids are stored in midgut cells and pseudoscorpions can subsist on these reserves for months after feeding. The short hindgut extends from the midgut to the anus at the tip of the anal cone.
Prey is captured with the pedipalps and then transferred to the chelicerae. In primitive pseudoscorpions such as Novobisium and Chthonius the chelicerae are used to masticate the food into a pulp as digestive fluid from the midgut is flooded onto it. The fluid is then sucked back into the gut leaving a dry masticated husk behind. In more derived groups the chelicerae tear a hole in the exoskeleton of the prey and digestive fluid is injected into the hole and then sucked back into the gut after the tissues are liquefied. The preoral cavity functions like a syringe to inject digestive fluid into the prey. The pharynx generates the negative pressure needed to suck fluid into the gut.
Figure 4. Diagrammatic sagittal section of a generalized pseudoscorpion. Redrawn from Weygoldt (1969). Pseudo13L.gif
Excretory System
Diverticula of the midgut form granules, thought to be guanine, which are released into the gut lumen and eliminated via the anus.
In addition there is a pair of coxal glands, which are end sac organs derived from metanephridia. The coxal glands open via nephridiopores between the coxae of legs 3 and 4.
Respiratory System
Pseudoscorpions rely on trachea for gas exchange. The two pairs of spiracles open laterally on the posterior margins of the sternites of abdominal segments 3 and 4 (Fig 2). The spiracles open into short tracheal trunks. Smaller branches radiate from the four trunks to supply the body.
Fluid Transport Systems
As very small animals pseudoscorpions rely heavily on diffusion to transport materials through the body. Most delivery is accomplished by the trachea (gasses) or by the digestive ceca (nutriment).
In addition a heart pumps blood through the hemocoel to supplement the other transport mechanisms (Fig 4, 18-33). The heart is a dorsal tube in segments 1-4 of the abdomen dorsal to the midgut. It bears 1, 2, or 4 pairs of ostia.
Nervous System
The nervous system is typically arachnid and strongly cephalized. The large two-part brain consists of protocerebrum and tritocerebrum filling the dorsal half of the cephalothorax (Fig 4, 18-33). The ventral cephalothorax is filled with the even larger subesophageal ganglion which comprises the coalesced segmental ganglia of all the segments of the cephalothorax and abdomen. The brain and subesophageal ganglion are connected laterally by connectives around the esophagus to form the circumesophageal nerve ring.
Pseudoscorpions usually have 2 or 4 eyes but occasionally have none. Most species probably have well developed chemoreceptive senses. The most important sensory system is the reception of vibrations. Long, specialized setae (trichobothria), especially on the pedipalps, are sensitive to very small movements of air. This is the chief sense used in locating and capturing prey. Pseudoscorpions can detect vibrations from prey from as far away as 15 mm, which is several times the body length of most pseudoscorpions.
Reproduction
There is typically sufficient sexual dimorphism for human observers to recognize the sexes. Females are usually larger than males. The female genital region is pale and only lightly sclerotized whereas that of the male is heavily sclerotized and much darker. Sometimes there are differences in the chelicerae and/or pedipalps.
The gonad is a single median tube in the abdomen lying between the midgut and its ventral diverticulum. It connects by a pair of gonoducts to the two gonopores (Fig 4, 18-33) located between sternites 2 and 3 on the anterior abdomen.
Sperm is transferred in pseudoscorpions via spermatophores. This is accomplished by a variety of behaviors (Fig 18-34). In general, the male presses his genital opening to the ground and extrudes a packet of sperm. He then raises himself high above the ground by extending the legs to draw the spermatophore away from the ground. This produces a spermatophore at the end of a long slender stalk. An interested female steps over the spermatophore and removes the sperm with her genital aperture.
In the simplest, most primitive, and most wasteful (of sperm) sperm transfer method, the male simply leaves his spermatophores scattered about the habitat whether or not females are present. Females may happen upon a spermatophore and engulf it but many spermatophores are never found are wasted. In more derived species, the male produces spermatophores only when females are present. In the most derived species the two sexes perform a complicated mating dance that insures that the spermatophore will have a willing recipient once it is produced.
Females brood the eggs and nourish them. The female constructs a brood nest of silk and other materials, lays eggs in it, and inhabits it until the first instar larvae hatch from the eggs. The eggs are poor in yolk reserves and the developing embryos are nourished by a fluid produced by the female.
There are three juvenile instars before the final molt produces an adult. Eggs hatch as a protonymph larva. The protonymphs molt to become deutonymphs and then tritonymphs. The next mole produces the adult. Molting ceases after the adult instar is reached.
The larvae are similar to the adults but are less heavily sclerotized and hence lighter in color. You may find some larvae in your leaf litter samples. They are smaller and paler than the adults. Larvae do not have the genital openings characteristic of the adults. The number and distribution of trichobothria and setae changes as the nymphs mature.
References
Gabbutt PD. 1965. The external morphology and life history of the pseudoscorpion Neobisium muscorum. Proc. Zool. Soc. London 145:335-358.
Hoff CC. 1958. List of the pseudoscorpions of North America north of Mexico.
Savory TH. 1977. Arachnida 2nd ed. Academic Press, New York. 340p.
Ruppert EE, Fox RS, Barnes RB. 2004. Invertebrate Zoology, A functional evolutionary approach, 7 th ed. Brooks Cole Thomson, Belmont CA. 963 pp.
Vanuytven H. Arachnology Home Page : www.arachnology.be/Arachnology.html
Warburton C. 1909. Arachnida, in Harmer, S.F. and A.E. Shipley eds, Cambridge Natural History vol IV Crustacea and Arachnids. MacMillan, London.
Weygoldt P. 1969. The Biology of Pseudoscorpions. Harvard, Cambridge. 145p.
Supplies
Dissecting microscope
Freshly collected or preserved pseudoscorpions
6-cm culture dish