Invertebrate Anatomy OnLine
Mercenaria mercenaria
Quahog
with notes on Tapes japonicus ©
24may2007
Copyright 2001 by
Richard Fox
Lander University
Preface
This is one of many exercises available from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises, a glossary, and chapters on supplies and laboratory techniques are also available at this site. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Mollusca P, Eumollusca, Conchifera, Ganglioneura, Ancyropoda, Bivalvia C, Metabranchia sC, Eulamellibranchia SO, Veneroida O, Veneroidea SF, Corbiculidae F (Fig 12-125, 12-122)
Mollusca P
Mollusca, the second largest metazoan taxon, consists of Aplacophora, Polyplacophora, Monoplacophora, Gastropoda, Cephalopoda, Bivalvia, and Scaphopoda. The typical mollusc has a calcareous shell, muscular foot, head with mouth and sense organs, and a visceral mass containing most of the gut, the heart, gonads, and kidney. Dorsally the body wall is the mantle and a fold of this body wall forms and encloses that all important molluscan chamber, the mantle cavity. The mantle cavity is filled with water or air and in it are located the gill(s), anus, nephridiopore(s) and gonopore(s). The coelom is reduced to small spaces including the pericardial cavity containing the heart and the gonocoel containing the gonad.
The well-developed hemal system consists of the heart and vessels leading to a spacious hemocoel in which most of the viscera are located. The kidneys are large metanephridia. The central nervous system is cephalized and tetraneurous. There is a tendency to concentrate ganglia in the circumenteric nerve ring from which arise four major longitudinal nerve cords.
Molluscs may be either gonochoric or hermaphroditic. Spiral cleavage produces a veliger larva in many taxa unless it is suppressed in favor of direct development or another larva. Molluscs arose in the sea and most remain there but molluscs have also colonized freshwater and terrestrial habitats.
Eumollusca
Eumollusca, the sister taxon of Aplacophora, includes all molluscs other than aplacophorans. The eumolluscan gut has digestive ceca which are lacking in aplacophorans, the gut is coiled, and a complex radular musculature is present.
Conchifera
Conchifera, the sister taxon of Polyplacophora, includes all Recent molluscs other than aplacophorans and chitons. The conchiferan shell consists of an outer proteinaceous periostracum underlain by calcareous layers and is a single piece (although in some it may appear to be divided into two valves). The mantle margins are divided into three folds.
Ganglioneura
Most Recent molluscs are ganglioneurans, only the small taxa Aplacophora, Polyplacophora, and Monoplacophora are excluded. Neuron cell bodies are localized in ganglia.
Ancyropoda
The mantle cavity, with its gills, is lateral. The calcareous portion of the shell is bivalve, with the valves opening laterally and joined dorsally by a derivative of the periostracum.
Bivalvia C
Bivalvia is a large, successful, and derived taxon. The body is laterally compressed and enclosed in a bivalve shell. The two valves are hinged dorsally. The the foot is large and adapted for digging in the ancestral condition. A crystalline style is usually present but never is there a radula. The mantle cavity is lateral and in most bivalves the gills are large and function in respiration and filter-feeding. The head is reduced and bears no special sense organs. The nervous system is not cephalized. The group includes scallops, clams, shipworms, coquinas, marine and freshwater mussels, oysters, cockles, zebra mussels, and many, many more.
Metabranchia sC
Most bivalves are metabranchs. The gills are adapted for filter feeding and water enters the mantle cavity posteriorly.
Eulamellibranchia SO
Eulamellibranchs have gills with tissue interfilamentar connections.
Veneroida O
Shell is usually equivalve and without a nacreous layer.
Laboratory Specimens
The clam Mercenaria mercenaria (= Venus mercenaria), variously known as the northern quahog (pronounced CO hawg), hardshell, littleneck, cherrystone, or chowder clam, is a common and commercially important species on the east coast of North America where it lives in soft sediments in shallow water (Fig 12-89, 12-92, 12-110F). This species is farmed commercially and is often available alive at reasonable cost in inland supermarkets and seafood markets. When thus available it offers a practical opportunity for study of the anatomy of a living bivalve, complete with beating cilia.
Preserved or living specimens of Mercenaria can be used for this study. The exercise is written for living specimens and these are recommended if available. Parenthetical comments refer to a similar species, the Japanese steamer clam, Tapes japonicus, which has been introduced to the west coast of North America and is available in seafood markets on the west coast.
The dissection should be conducted in isotonic magnesium chloride (if living) or tapwater (if preserved. Use the dissecting microscope as needed.
External Anatomy
Shell
Study a complete, dried, empty shell. The thick chalky shell of Mercenaria consists of two similar valves, which fit tightly together to enclose and protect the soft parts of the animal (Fig 1, 12-92B). (The shell of Tapes is neither thick nor chalky). The exterior of each valve is rough and ornamented with irregularly spaced, raised, concentric growth ridges. (Tapes has, in addition, distinct radial ridges.)
Figure 1. Oblique view of the right side of an intact shell of Mercenaria mercenaria. Bivalve63L.gif
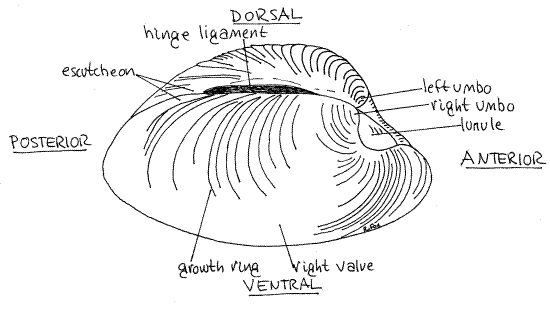
The shell is bilaterally symmetrical and the plane of symmetry passes through the hinge to divide the shell into right and left valves. The valves are nearly identical in size and shape, a condition referred to as equivalve. Some bivalves, oysters for example, have right and left valves of different size and shape and are inequivalve.
Each valve is strongly convex outwards and bears a conspicuous raised protuberance, the umbo (= beak), on its dorsal surface near the hinge (Fig 1, 2). The umbo is the oldest part of the valve and the concentric growth ridges are centered around it. When the valves are closed, the umbos almost touch each other on the midline. The umbos are dorsal and slightly anterior to the middle of the hinge.
The two valves are joined dorsally at the flexible (in life) hinge and are free to separate from each other along the ventral margin. The dark brown, proteinaceous hinge ligamentoccupies part of the hinge (Fig 1, 2, 12-92B). The hinge ligament is a good dorsal landmark. The small (in life) gap between the ventral margins of the opened valves is the gape. The gape of dead specimens is far wider than in life.
Use the hinge and umbos as landmarks to recognize dorsal and anterior, respectively, and, having accomplished that, decide which valve is left and which is right. Find posteriorand ventral and identify the plane of symmetry.
Figure 2. Interior of the left valve of the shell of Mercenaria mercenaria. Bivalve64La.gif
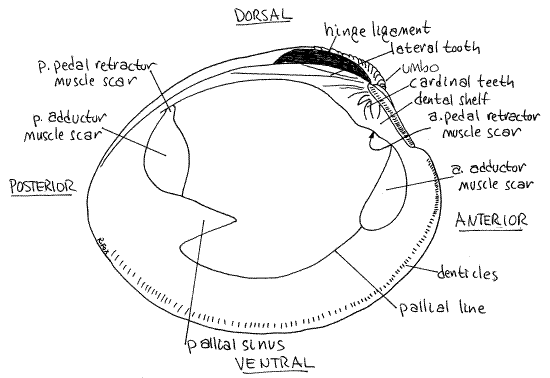
Place the two valves together and look at the dorsal surface. The most obvious features are the umbos. Anterior to the umbos is a conspicuous, heart-shaped depression known as thelunule (Fig 1). It is a characteristic feature of Veneridae. Posterior to the umbos is a broad, flat, indistinctly defined platform, the escutcheon. (The escutcheon is small and obscure inTapes.) The free edge of the posterior right side of the escutcheon overlaps the free edge of the left so that, even when the clam is open, this part of the gape is protected, perhaps from falling sand grains. (The overlap in Tapes is very slight.)
Viewed from the side, each valve is lopsided, with a longer bluntly pointed posterior end and a shorter, smoothly rounded anterior end. (The posterior end of Tapes is not pointed.) Valves with asymmetrical anterior and posterior halves are said to be inequilateral. Mercenaria and Tapes are only moderately inequilateral, mussels are strongly inequilateral.
Look at the medial side of one of the two valves (Fig 2, 12-92B). Dorsally, the margin is occupied by the hinge, which is the articulation between the two valves. The hinge ligament is a conspicuous feature of the hinge. The ligament is composed of protein (conchiolin) and may or may not be present on dry empty shells. Look at a fresh, undissected specimen and find the ligament if your empty shell does not have one. The ligament of living animals is elastic but that of dry valves is hard and brittle.
The hinge is equipped with strong teeth to assure the proper alignment between the valves each time the shell closes. The teeth are located on a platform called the dental shelf (Fig 2). Most conspicuous of these are the short, vertical, or oblique cardinal teeth located directly ventral to the tip of the umbo. Each valve has three cardinal teeth but one tooth on the left is obscure making it appear as if there are only two teeth on this side. (In Tapes all three teeth are distinct.) A posterior lateral tooth is located posterior to the cardinal teeth. It is a long, low, rough, irregular ridge with its long axis oriented anterior to posterior. Unlike the cardinal teeth it does not look like a tooth.
The anterior and ventral margins of the valves bear fine teeth known as denticles (Fig 2). (Tapes lacks denticles.) The denticles probably prevent shear (sideways slipping) when the valves are pulled tightly together by the powerful adductor muscles. Hold the two valves together as they would be in a living clam with its valves closed. If you have the cardinal teeth properly aligned, the valves will fit snugly together and cannot be made to slip past each other.
The large, smooth anterior and posterior adductor muscle scars are easily seen at their respective ends of the inside of each valve (Fig 2, 12-92B). In intact clams the two adductor muscles run transversely from one valve to the other and their action is to pull the valves together (adduct) and hold them closed. Two additional muscles, the anterior and posterior pedal retractor muscles, insert on the shell near the adductor muscles. The anterior pedal retractor scar is separated by a small space from the anterior adductor scar and is under the anterior end of the dental shelf. (The anterior pedal retractor muscle scar is easy to see in Tapes and is located just anterior to the dental shelf.) The posterior pedal retractor scar is continuous with the posterior adductor scar (Fig 2).
The pallial line runs from one adductor to the other and parallels the ventral margin of the valve. It is the site of insertion of pallial muscles in the mantle and is indented sharply beside the posterior muscle scar. This V-shaped indentation is the pallial sinus and is a recess in the line of pallial muscles for the withdrawn siphons. (“Pallial” is another word for mantle.)
The inside of each valve of M. mercenaria is white with posterior purple markings. The shells of this species were cut into beads and used by east coast North American Indians as money, called wampum. This is the basis of the scientific name "mercenaria" (mercenari = hired for wages).
The typical mollusc shell consists of three layers (Fig 12-91). The outermost is the periostracum composed of the protein conchiolin. This layer is thin, eroded, and insignificant inMercenaria. The dull, chalky prismatic layer (ostracum) is the middle layer. It is exposed on the outside of the valve due to the absence of the periostracum. The innermost layer, and the one in contact with the clam, is the calcareous lamellar layer (hypostracum) which is smooth and glossy. (The thin, brown periostracum of Tapes remains intact and the prismatic layer is not evident.)
Soft Anatomy
" Open the shell of your clam to study the soft anatomy within. The adductor muscles keep the shell closed and it cannot be opened until they are severed. If you are dissecting a preserved specimen, the valves may already be pegged open with a wooden wedge. If this is the case, you may skip the remainder of this paragraph and proceed to the instructions for cutting the adductor muscles.
If you have a living specimen or an unpegged preserved specimen, you must make anterior and posterior openings in the shell to give you access to the adductor muscles so you can cut them. This is not easy to do but the best procedure is to use a pair of pliers to pinch away the anterior and posterior edges of the shell. Do not strike the shell as it cracks easily. First refer to the empty valves to help determine the positions of the two adductor muscles and begin removing the shell closest to them.
Gain access to the anterior adductor first by pinching away the anterior edges of the contiguous valves. You will not be able to get as close to the posterior adductor muscle as to the anterior because the posterior shell does not provide a raised edge to pinch. Remove enough shell to create a narrow opening at each end.
Insert a scalpel blade into the anterior opening and feel for the adductor muscle. Use the longest and sharpest blade you have but be very careful that you do not cut anything until you find the muscle. The muscle is firm, rubbery, and unyielding whereas the other tissues are soft and do not resist gentle probing. The muscle is easy to recognize by touch. Once you have found the muscle, sever it completely with the scalpel. Do the same with the posterior adductor muscle. The pedal retractor muscles should be cut also.
Arrange the clam so the left valve is up, facing you, and you are looking at the left side of the animal. With all the muscles severed, carefully lift the left valve a little so you can see into the gape. The body will be cradled in the concavity of the right valve but a thin sheet of tissue, the left mantle skirt, adheres closely to the inner surface of the left valve. Slip the blunt end of your scalpel handle between the left mantle skirt and the left valve and use it to free the soft tissue from the shell. Carefully work the scalpel handle around the stumps of the adductor muscles being careful that you do not damage the mantle. Note the two pedal retractor muscles that insert on the shell beside the much larger adductor muscles. Cut them now if you did not do so earlier. When the mantle is free, detach the left valve at the hinge and set it aside.
Place the clam in a culture dish or dissecting pan with the right valve down. Cover the clam with isotonic magnesium chloride (if living) or tapwater (if preserved).
>1a. Look at the inner surface of the detached left valve for a difference in the appearance of the lamellar layer proximal and distal to the pallial line. The surface of the lamellar layer inside the pallial line, is alternately eroded and then redeposited as part of the normal metabolic operations of the animal. When the valves are closed and the clam is forced to respire anaerobically, calcium is removed from this region to neutralize the succinic acid produced by this metabolism and calcium succinate is produced. When the valves open and the clam switches to aerobic respiration the calcium is redeposited. <
Muscles
Look at the left side of the clam and find the anterior and posterior adductor muscles (Fig 3). The muscle fibers run transversely, across the clam, from valve to valve and their action is to adduct, or close, the valves.
Each muscle is composed of two distinct regions (Fig 3). The anterior part of the anterior adductor muscle and the posterior part of the posterior adductor muscle are catch musclescomposed of smooth muscle fibers with abundant connective tissue. They are capable of sustained, slow contraction and in life are white. The remainder of each muscle is the fast musclecomposed of obliquely striated fibers with abundant supplies of myoglobin and mitochondria but with relatively little connective tissue. In life fast muscle is red due to its myoglobin. Fast muscle contracts and relaxes rapidly but fatigues easily and is incapable of sustained contraction. The adductor muscles are opposed by the elastic recoil of the hinge ligament.
The pedal retractor muscles are paired. Find one of the small anterior pedal retractor muscles (Fig 3). It is a short cord of muscle and connective tissue running from the anterior end of the foot to the shell. The posterior pedal retractor muscles are similar cords extending from the posterior end of the foot to the shell. The retractors pull the foot back into the shell as is necessary before closing the gape, or, when digging, to pull the clam toward an expanded foot anchored in the sand. Mercenaria lack a pedal protractor muscle.
Mercenaria exemplifies the dimyarian condition in having two similar adductor muscles. Many bivalves, notably scallops and oysters, are monomyarian and have only one adductor muscle (the posterior). Others, such as mussels and pens, are heteromyarian and have a large posterior muscle and a reduced anterior muscle.
Bivalves do not have abductor muscles to open the valves. The adductor muscles extend across the animal from one valve to the other. When they contract they pull the valves together, close the gape, and stretch the elastic hinge ligament. When the muscles relax, the ligament returns to its original resting shape and in doing so pulls the dorsal ends of the valves toward the midline (toward each other). This, of course, means that the ventral margins of the valves move away from each other, thereby opening the shell and creating a narrow gape through which the foot can be extended.
The ligament of Mercenaria is outside the hinge. Some bivalves also have an internal ligament (= resilium) on the inside of the hinge that is compressed, rather than stretched, when the valves close. When the muscles relax, the elastic resilium pushes the valves apart, and this also opens the gape. Mercenaria does not have a resilium.
Mantle Skirts
The bivalve body is enclosed on the right and left by two large, thin double layers of body wall (Fig 5). These are the right and left mantle skirts. Each is a thin sheet of tissue, free at the ventral edge but connected with the body dorsally. The pallial muscles attach the mantle skirt to the shell along the pallial line.
Mantle Cavity
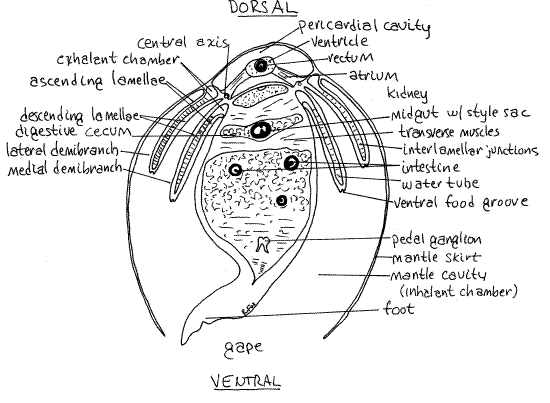
Between the right and left mantle skirts lies a large space, the mantle cavity, in which lies the body (Fig 3, 5). The mantle cavity is continuous with the sea and is filled with seawater. Without cutting tissue, lift the left mantle skirt and find the mantle cavity. It is divided into a large ventral region below and beside the body, known as the inhalant chamber (= branchial chamber), and a smaller dorsal region above the body, the exhalant chamber (= suprabranchial chamber) (Fig 5, 12-90). The two chambers are separated from each other by the gills. The space you see now is the spacious inhalant chamber and you cannot yet see the much smaller exhalant chamber.
Look into the mantle cavity (inhalant chamber) and make a quick identification of the most conspicuous features of the body so you can use them for landmarks later. With the left mantle skirt held out of the way, closest to you is the long, leaflike left gill (Fig 3, 5). Medial to the gill is the large bulging visceral mass with the thinner, muscular, bladelike (in life) foot attached to its ventral midline. Lift the foot and visceral mass to see the right mantle cavity, right gill, right mantle skirt, and right valve, in that order.
Mantle Folds
Study the ventral margin of the right mantle skirt with magnification. The mantle margin of Mercenaria and other venereid clams is composed of four folds although most bivalves have only three (Fig 4, 12-91). The homologies between the four venerid folds and the three folds of other bivalves are unclear but the extra fold probably arose through subdivision of the inner fold.
Figure 3. The left side of Mercenaria with the left valve and left mantle skirt removed. Bivalve65La.gif
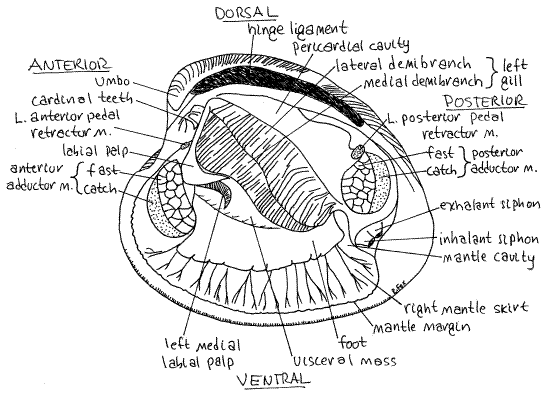
The outer fold, which is adjacent to the shell, secretes the prismatic layer and periostracum (Fig 4, 12-91). The lateral side of the outer fold secretes the prismatic layer of the shell whereas the medial side secretes the periostracum. The lamellar layer is secreted by the entire lateral surface of the mantle skirt.
The inner fold is muscular and in Mercenaria is double. The wide border of the mantle, from its free edge to the pallial line, is thicker than the rest of the mantle due to the presence of the pallial muscles that insert on the shell along the pallial line. These muscles can be seen radiating across the mantle border peripheral to the pallial line. Tug gently on the free edge of the right mantle and observe that (in life) it is firmly attached to the shell along the pallial line.
The middle fold lies between the inner and outer folds and is sensory. It is weakly developed in Mercanaria.
The groove between the outer and middle folds is the periostracal groove. In living specimens the freshly secreted periostracum can be seen extending out of this groove across the exposed surface of the margin of the valve. It is a very thin, transparent, delicate, glistening membrane.
Siphons
Posteriorly, the right and left mantle skirts coalesce with each other across the midline to form the two short tubular siphons (Fig 3, 12-89). The siphons are darkly pigmented and each has short sensory tentacles surrounding its outer aperture. The ventral one is the inhalant siphon and the exhalant siphon is dorsal. These short tubes are formed by the fused right and left posterior mantle skirts. (In Tapes the siphons are long tubes and are pigmented only at their tips.) The siphons are elaborations of the fused right and left mantle margins. The sensory tentacles are part of the middle mantle fold, which is the sensory fold. In its life position Mercenaria is shallowly embedded in sediment with only the tips of the short siphons exposed above the sediment surface. It is appropriate that the greatest concentration of sense organs is on the siphons rather than around the periphery of the gape. What would you predict about the life position of Tapes?
The inhalant siphon brings water into the inhalant chamber. Insert your blunt probe into the external opening of the inhalent siphon and show that water entering here would enter the inhalant chamber of the mantle cavity. Water in the inhalant chamber passes through tiny openings, the ostia, in the gills to enter the exhalant chamber, from which it then exits via the exhalent siphon (Fig 12-89B).
A delicate fold of tissue, located dorsal to the inner aperture of the inhalant siphon and known as the siphonal membrane, can be extended from its retracted resting position to partially cover the inner aperture of the inhalent siphon. This deflects water that normally would have gone directly to the gill surfaces, to the ventral inhalant chamber where it stirs and resuspends the accumulated debris and pseudofeces so they can be more easily expelled by the next contraction of the adductor muscles. The siphonal membrane is easily seen on the dorsal surface of the inner aperture of the inhalant siphon.
Dorsal to the siphons the entire right and left mantle margins are fused to each other on the midline beneath the hinge. A special secretory region of the dorsal mantle, ventral to the ligament, is called the subligamental ridge. It secretes the ligament.
Gills
" With scissors, remove the left mantle skirt and look at the left gill. Each gill consists of a central axis from which are suspended sheets of fused filaments. On each side is a single complete gill, or holobranch (Figs 3, 5, 12-96D). A holobranch is composed of two delicate, leaflike demibranchs, or half gills. These are the outer lateral demibranch and the innermedial demibranch (Figs 3, 5, 12-90). The two are joined dorsally along the central axis which is attached to the body (Fig 5, 12-90).
Each of the two surfaces of a demibranch is a lamella (Fig 5). One surface is the descending lamella firmly attached to the central axis and the other is the ascending lamella weakly attached to either the mantle or the visceral mass (Fig 5). In Mercenaria, but not Tapes, the connections of the ascending lamellae to the body are fragile and the margins of the gills are easily torn free of their attachments. At present you are probably looking at the ascending lamella of the lateral demibranch of the left gill.
Figure 4. Cross section through the ventral edge of a valve and the associated mantle skirt of Mercenaria. Bivalve66L.gif
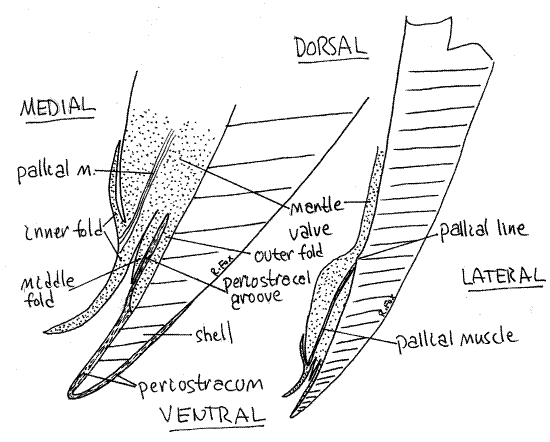
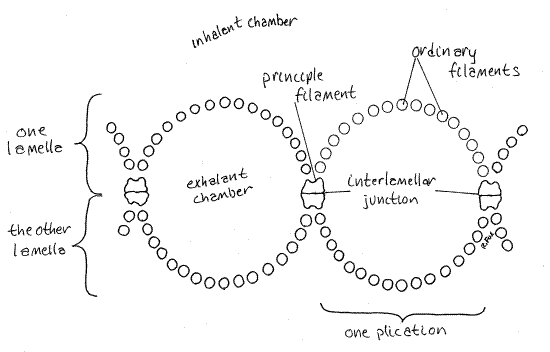
The surface of the lamella is conspicuously ridged. Each ridge is a plica, running parallel to the short axis of the gill. Each plica is made up of several gill filaments and contains a water tube (Fig 5, 12-98C,D). The water tubes are part of the exhalant chamber.
At regular intervals a thicker filament is specialized to form the margins of the water tube, separate it from adjacent water tubes, and hold the opposite lamellae together. These special filaments are larger are the "principle filaments". The other, more numerous and smaller filaments are known as "ordinary filaments". Two successive principle filaments and the ordinary filaments between them form one plica, or ridge. The opposite principle filaments are held together by permanent interlamellar junctions.
Figure 5. Cross section of Mercenaria with the shell omitted. Bivalve67.La.gif
Examine one of the plicae with the highest power of your dissecting microscope and you will see faintly the long slender gill filaments that make up its walls. Venus clams such asMercenaria and Tapes, have eulamellibranch gills in which adjacent filaments are permanently attached to each other by broad tissue interfilamentar junctions (Fig 12-98). The only openings between filaments are the microscopic ostia. If you are having trouble making out the details of the gill surface because of poor contrast, you may be able to improve the situation by placing a little 1% toluidine blue/seawater on the surface of the gill.
The opposite walls of each water tube are made of opposite ends of the same filaments (Fig 12-98C,D). The medial surface (lamella) of the lateral demibranch is composed of the proximal (descending) end of the filaments and the lateral surface of the distal (ascending) end of the same filaments. The connections from one lamella to the other are interlamellar junctions and they contain blood vessels.
The ciliated longitudinal groove along the ventral edge of each demibranch is a ventral food groove used to transport food particles anteriorly to the mouth (Fig 5). The ciliary beat in the food grooves is anterior, toward the head, labial palps, and mouth whereas that of the lamella is either dorsal or ventral.
Figure 6. Frontal section of two plications (vertical channels) of a scallop gill. Bivalve84La.gif
During feeding and respiration a water current bearing food and oxygen enters the inhalant chamber via the inhalant siphon. The water enters the ostia and passes into the water tubes of the exhalant chamber. Food particles are filtered from the current as it passes through the ostia, remain on the inhalant side where they are mixed with mucus and transported to the food grooves by cilia.
>1b. If your clam is living, remove it from its dish and place it, on a cloth towel, on the stage of your dissecting microscope. Do not get seawater or magnesium chloride on the microscope. Arrange the animal so the surface of a lamella is horizontal. Place a drop of carmine/seawater suspension on the lamella. Watch the motion of the particles along the face of the lamella and in the ventral food groove. <
>1c. With scissors, remove a small square (about 5x5 mm) from the lateral demibranch, being sure to include the ventral food groove. Make a wetmount and examine it with the compound microscope. Find the food groove along one margin and use it as a landmark. Adjust the light, focus carefully on the uppermost of the two lamellae, and study the construction of the demibranch. The surface of the lamella is composed of large parallel ridges, the plicae, which are themselves formed of smaller ridges, the gill filaments. Focus down to bring the filaments of the other lamella into view. Trace a filament to the food groove and note that it reverses direction here and extends up the opposite side of the demibranch. Look in the grooves between the filaments and you may be able to see the small oval ostia.
Labial Palps
The long narrow labial palp on each side of the mouth is an anterior extension of the gill (Figs 3, 7). Each palp consists of a lateral and a medial lamella. The lateral demibranch of the gill connects with the lateral lamella of the palp and the medial demibranch with the medial lamella. Each lamella bears a sorting field of ciliated ridges and grooves on one surface (Fig 12-100). The fields are arranged so that when the lamellae lie together, the sorting fields face each other. A ciliated oral groove lies between the tow sorting fields and leads to the mouth.
Mucus and food particles from the gills move along the food grooves to the labial palps where the sorting fields separate desirable organic matter (food) from indigestible mineral particles. Organic particles and mucus move via the ciliated oral groove to the mouth whereas mineral particles, also bound in mucus, move to the tip of the palp from which they are dropped into the inhalant chamber. This mixture of discarded mineral particles and mucus is known as pseudofeces and it is periodically expelled through the inhalant siphon by sudden contractions of the adductor muscles.
>1d. If you have a living specimen, place carmine/seawater on the sorting field of one of the labial palps to visualize the ciliary currents. Look for particles moving parallel to the ridges and others moving across the top of the ridges but perpendicular to them. Look also for a string of particles and mucus moving along the margin to the distal tip of the lamella and still another moving along the oral groove between the lamellae to the mouth. You may be able to see the food string disappear into the mouth. Repeat with chalk dust collected from the chalk tray of a blackboard. <
Trace the lamellae of the labial palps across the midline and note that each is connected with its counterpart on the opposite side of the body. These two thin folds of tissue are the lips. The lip that connects the two lateral palps is the upper lip and the one that connects the two medial palps is the lower lip (Fig 12-100). The oral groove is between them. Exactly on the midline, between the upper and lower lips, is the mouth. Tilt the animal and find the mouth with the dissecting microscope. This area may have been damaged when the anterior adductor muscle was cut. The mouth is easiest to find if it has a string of carmine red mucus entering it.
Internal Anatomy
Hemal System
The bivalve hemal system consists of heart, arteries, blood, and an extensive hemocoel. Only the heart will be studied in this exercise.
The heart lies in the pericardial cavity located dorsally just below the hinge (Figs 3, 5, 7, 12-89B). It is surrounded by the thin membranous pericardium. The body wall in this area is very thin and the pericardium is close to the surface. The pericardial cavity is a remnant of the coelom and the pericardium is peritoneum. The dark reddish brown (pale beige in Tapes)nephridium, or kidney, can be seen on the walls of the pericardial cavity, especially posteriorly. Paler, greenish brown tissue situated farther anterior is the digestive cecum.
" With fine scissors, make a shallow, longitudinal, middorsal incision in the thin dorsal body wall and pericardium paralleling and extending the length of the hinge ligament. This cut opens the pericardial cavity to reveal the heart within. The heart consists of a single large ventricle into which the paired right and left atria empty (Fig 7). The ventricle lies on the midline and is a long, soft, ovoid organ. It is penetrated by the posterior intestine, or rectum, which runs the length of its lumen (Fig 7). The intestine can be seen faintly through the walls of the ventricle.
Figure 7. Dissection of the left side of Mercenaria. The left side of the visceral mass has been dissected. The left mantle skirt and most of the kidney and left gill have been removed. Bivalve68La.gif
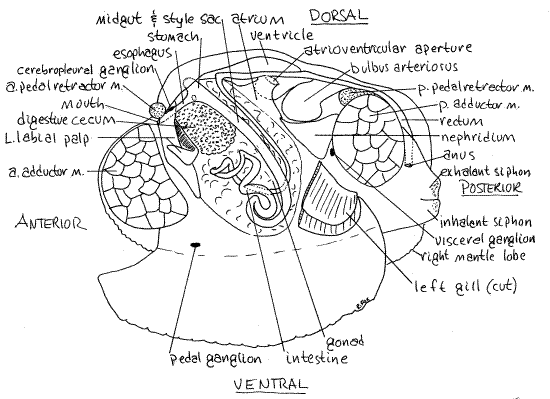
With a blunt probe, carefully lift the edge of the ventricle and find the left atrium attached to its left side (Fig 7). The atrium is a thin-walled, transparent, truncated triangle extending from the ventrolateral pericardial wall to the ventrolateral wall of the ventricle. The truncated apex of the atrium attaches to the ventricle and its broad base extends along the length of the central axis of the gill. Oxygenated blood from the gills drains into the atria and from there enters the ventricle.
>1e. If your animal is alive, count the heartbeats for three minutes and calculate a pulse rate for your specimen. If your animal is in magnesium chloride it will have to be transferred to seawater before the heart will beat. <
Anteriorly, the ventricle narrows to become the wide, transparent anterior aorta which may be visible beside the base of the anterior pedal retractor muscle. The anterior aorta is dorsal to the intestine. The intestine emerges from the visceral mass at the end of the aorta and bends almost 90° to enter the lumen of the ventricle (Fig 7).
Posteriorly, the ventricle constricts to become the posterior aorta which then immediately expands to form the large swollen bulbus arteriosus. The size of the bulbus varies depending on the amount of blood it contains. The posterior aorta then narrows and continues posteriorly, ventral to the rectum.
>1f. With fine scissors, open the ventricle with a longitudinal middorsal incision. The rectum, which runs through the lumen of the ventricle, is easily seen. Use fine forceps to pull the cut edge of the ventricle toward you so you can look inside and see the inner surface of the left ventricular wall. Find the atrioventricular aperture between the atrium and ventricle and insert a probe into it to demonstrate continuity between the two chambers. Notice the bundles of muscle fibers in the lumen of the ventricle. <
Exhalant Chamber
The ventrolateral floor of the pericardial cavity is the roof of the exhalant chambers (Fig 5). Insert the blunt probe into the exhalent aperture and into the exhalant chamber. The transparent roof of the chamber extends anteriorly from the exhalent siphon but may have been cut when you severed the posterior adductor muscle. The visceral ganglion of the nervous system is in the tissue between the roof of the chamber and the adductor muscle. In living clams the ganglion is orange (neuroglobin). Be careful that you do not destroy it.
>1g. Open the exhalant chamber by tearing the ascending lamella of the lateral demibranch away from its attachment to the mantle. Look inside the chamber and find the upper ends of the water tubes inside the plicae. Insert a blunt probe into the exhalent siphon to demonstrate its continuity with the exhalant chamber. Above the gills the exhalant chamber is divided into two longitudinal channels, one above each demibranch. The attachment of the central axis of the gill to the dorsal mantle wall separates the two chambers. The medial exhalant chamber lies above the medial demibranch and the lateral exhalant chamber lies above the lateral demibranch. Look at the cut gills and find water tubes and the interlamellar junctions that define them. Much of the roof of the exhalant chamber is also the floor of the pericardial cavity. The dark nephridium can be seen in the roof of the chamber at the posterior end of the pericardial cavity. Another dark portion of the nephridium can be seen at the anterior end of the pericardial cavity. <
Excretory System
The two nephridia , or kidneys, are large metanephridia, one on each side on the floor and walls of the pericardial cavity at the dorsal margin of the visceral mass (Figs 5, 7, 12-89B). They are brown or reddish-brown (in life) and are easily seen without dissection although their structure is not apparent. The nephridia are elaborate ducts extending from the pericardial cavity to nephridiopores in the exhalant chamber. Urine is released from this pore into the exhalant water stream.
>1h. You may want to study the nephridium in more detail (but a freshwater mussel would be a better choice for this purpose). The nephrostome is a small opening from the antero-ventral corner of the pericardial cavity into anterior end of the nephridium. Most of the inner wall of the pericardial cavity is smooth but where it covers the region of the nephrostome it is folded minutely and the nephrostome is hidden in the folds. Sometimes it helps to put a little pressure on the adjacent nephridium to cause fluid to be extruded from the opening. The nephrostome opens into the renopericardial canal that leads into the lumen of the nephridium. The nephrostome is a ciliated funnel and the nephridium lumen is derived from coelomic space. The antero-ventral region of the nephridium, into which the canal opens, is glandular. The lumen of this region is continuous with a thin-walled postero-dorsal region that functions as a reservoir, or bladder. The right and left nephridia are connected across the body by a small duct. Each nephridium extends posteriorly from the nephrostome, mostly embedded in other tissues. The reservoir opens into the exhalant chamber via a small nephridiopore. The nephridiopore is located in a tiny dimple on the roof of the exhalant chamber. This inconspicuous pore is between the medial demibranch and the visceral mass and is at about the level of the posterior end of the ventricle. It is just posterior to the small genital papilla on which the gonoduct opens. <
Digestive System
The bivalve gut consists of a mouth, esophagus, stomach, digestive ceca, midgut, rectum, and anus. A complex stomach with style and style sac are present but bivalves have no buccal cavity, radula, radular sac, or odontophore. Most of the gut is within the visceral mass and cannot be seen without dissection. If you plan to study the nervous system, refer to the nervous system section and find the cerebropleural ganglia at this time as they are usually destroyed during the dissection of the anterior gut.
" Remove the left gill and left labial palp so you have unobstructed access to the left side of the visceral mass. Relocate the mouth on the anterior midline between the upper and lower lips of the labial palps (Fig 7, 12-89B). The mouth opens directly into the esophagus. The short esophagus passes through the greenish digestive cecum to join the stomach (Fig 7). The anterior end of the stomach, which is otherwise embedded in the visceral mass, lies very close to the surface of the antero-dorsal corner of the visceral mass and can sometimes be seen without cutting.
" Insert a blunt probe through the mouth into the esophagus to the stomach. The probe will follow the gut lumen and you can see it (the probe) through the gut wall. The esophagusextends obliquely dorso-posteriorly between the bases of the two anterior pedal retractor muscles (Fig 7). The esophagus more or less parallels the edge of the dental shelf in the vicinity of the cardinal teeth. The walls of the esophagus bear fine, longitudinal folds, or rugae.
Open the stomach by inserting a blade of your fine scissors into the mouth and cut along the esophagus, following the probe to the stomach. Once you have entered the stomach, cut posteriorly through the lateral wall of the visceral mass and digestive cecum, opening the stomach as you go (Fig 7). Continue this cut posteriorly, paralleling the dorsal border of the visceral mass. The incision should follow and open the intestine (= midgut) which extends posteriorly from the stomach. Continue this cut, following the intestine, around the posterior curve of the visceral mass to the foot.
Periodically use a Pasteur pipet or squirt bottle to blow away the accumulating debris that would otherwise obscure your view of the gut lumen. Examine the intact right wall, floor, and roof of the stomach (Fig 12-102). Find the opening of the esophagus on the midline at the anterior end of the stomach. It lies under a fleshy overhang of tissue.
Two openings to the digestive ceca are located on the floor of the stomach just posterior to the opening of the esophagus. A branching network of tubules extends from these openings throughout the ceca (Fig 12-103B).
On the right of the esophageal opening, and extending dorsally to the left side, is a large, ciliated sorting field (Fig 12-102). The field is composed of numerous fine ridges and grooves. A thick shelf on the right side of the stomach marks the posterior right border of the sorting field and separates the dorsal and ventral portions of the stomach from each other.
The roof and much of the left wall of the stomach are covered by a thin, chitinous gastric shield but it is difficult to see (Fig 12-102). Although the shield itself is nearly invisible, the wall of the stomach under it is smooth and can thereby be distinguished from the surrounding walls.
The intestine extends from stomach to anus (Fig 7). Its anterior end is the midgut whereas the rectum is its posterior end. The midgut consists of a descending intestine and ascending intestine. The descending intestine exits the posterior end of the stomach and parallels the dorsal and posterior edges of the visceral mass. It is divided into two parallel tubes by two longitudinal ridges of tissue that approach each other in the middle of the midgut lumen (Fig 5, 12-103A). These two ridges are typhlosoles. The typhlosoles are ciliated ridges that transport particulate material.
The major typhlosole is on the lower right side of the midgut and is the larger of the two. It begins in the anterior stomach and its edge is thin and sharp. The minor typhlosole begins at the posterior end of the stomach, lies on the dorsal left wall of the midgut, and has a broad, bilobed edge.
The two side by side tubes of this region of the gut are the dorsal style sac and the ventral intestine. The style sac of your specimen may or may not contain a crystalline style. The style is produced only when needed and if your animal has been maintained without food, it probably will not have a style. The style is a very long, narrow, flexible, pellucid, gelatinous rod that occupies the length of the style sac and protrudes from the sac into the stomach lumen. It is composed of hydrolytic enzymes (amylase, cellulase, lipase) secreted by the glandular walls of the sac. It stirs the stomach contents, serves as a windlass to help pull mucus/food strings from the labial palps into the stomach, and is a source of digestive enzymes for the extracellular hydrolysis of carbohydrates and lipids in the stomach. Protein digestion occurs intracellularly in the digestive ceca.
The intestine extends posteriorly from the stomach, curves around the posterior edge of the visceral mass to the foot where it changes direction and turns anteriorly as the ascending intestine (Fig 7). It soon changes direction again and heads dorsally. Only the intestine, and not the style sac or the style, continues past this point.
The intestine loops in a characteristic pattern through the visceral mass, and emerges at the base of the anterior aorta (Fig 7). Here it makes a right angle turn to extend posteriorly through the center of the pericardial cavity and ventricle as the rectum. Beginning at the point where the style sac ends, the intestine is characterized by the presence of a single, large, roundedtyphlosole that occupies most of its lumen (Fig 5). Continue tracing the intestine a little beyond the point where the typhlosole begins so you can get a good look at this part of the gut. The gut is easiest to trace by opening it and following the typhlosole. Trace the intestine as far as you wish into the visceral mass. If you have the time you may follow it all the way to the heart but the procedure is tedious and time-consuming.
Locate the rectum where it exits the heart and trace it posteriorly. It passes between the bases of the two posterior pedal retractor muscles and then curves over the top of the posterior adductor muscle and part of the way around its posterior margin. It ends in the exhalant chamber at the anus on the posterior edge of the adductor muscle. The anus is dorsal to the inner aperture of the exhalent siphon. Fecal pellets released from the anus are caught in the exhalant current and swept out the siphon. (In Tapes, the rectum flares trumpetlike to end at the anus.)
Nervous System
The bivalve nervous system is relatively simple and consists of four major pairs of ganglia (cerebral, pleural, pedal, and visceral), connectives and commissures between them, and the nerves radiating from them (Fig 12-119).
Despite its simplicity, the system is not easily studied. The ganglia can usually be found but tracing the connectives and nerves is difficult. In living animals the ganglia are orange or yellow with neuroglobin, making them easier to recognize. The bivalve sensory system is weakly developed. Most sensory receptors are in the middle fold of the mantle margin.
The cerebral and pleural ganglia are coalesced and the resulting two cerebropleural ganglia are close to each other atop the anterior end of the esophagus, very close to the mouth (Fig 7). They are embedded in connective tissue on the posterior dorsal edge of the anterior adductor muscle. The two cerebropleural ganglia are connected to each other via a short transverse cerebral commissure across the top of the esophagus and nerves extend from them to the labial palps, anterior adductor muscle, anterior mantle, and anterior pedal retractor muscle.
From the cerebropleural ganglia a pair of visceral nerves (= cerebrovisceral connectives) runs posteriorly through the dorsal visceral mass to the visceral ganglia. The visceral ganglia are located on the anterior face of the posterior adductor muscle between the origins of the two posterior pedal retractor muscles (Fig 7). They are fused with each other on the midline. The visceral ganglia send nerves to the siphons, posterior adductor muscle, posterior pedal retractor muscle, posterior mantle, nephridium, gill, and heart.
A pair of cerebropedal connectives extends ventrally and posteriorly from the cerebropleural ganglia to the pedal ganglia in the foot. The two pedal ganglia are fused with each other and lie on the midline in the visceral mass, very near the muscles of the foot (Figs 5, 7). The pedal ganglia innervate the musculature of the foot. Look for them on the border between the foot and the visceral mass ventral and anterior to the point at which the midgut makes its sharp turn dorsally.
Reproductive System
Like most bivalves, Mercenaria is gonochoric and the two gonads, either ovaries or testes, fill most of the space in the visceral mass (Figs 5,7). The size of the gonads depends on the degree of development. Although there are two of them, they cannot usually be distinguished from each other.
>1i. Determine the sex of your clam by making a wetmount of some of the gonadal tissue and examining it with the compound microscope. Look for gametes, either large sphericaleggs or tiny flagellated spermatozoa. <
The gonads occupy remnants of the embryonic coelomic cavity but that will not be apparent in gross dissection. Each gonad empties into the exhalant chamber via a gonoduct that opens at a gonopore at the tip of a small genital papilla just anterior to the nephridiopore. The genital papilla is located at about the level of the posterior end of the ventricle in the medial exhalant chamber but is difficult to locate.
Cross Sections
>1j. If extra specimens are available, you may want to make a cross section. This works best if the tissues are firm, as are those of preserved or steamed specimens. If you have fresh or living specimens, steam them, shell and all, in a pot with a little boiling water for about ten minutes to denature and solidify the proteins. Remove the clam from the shell and, using a very sharp scalpel, razor blade, or scissors make one or more cross sections entirely through the body including the gills and mantle. Most useful is a slightly oblique section passing through the heart and anterior end of the base of the foot (Fig 5). Rinse the sections gently and then immerse them in fluid in a small dissecting pan for study with the dissecting microscope. Use Figure 5 to identify the structures already familiar to you from your conventional dissection. <
References
Brooks WK . 1890. Handbook of Invertebrate Zoology. Bradlee Whidden, Boston. 352p.
Brown FA . (ed) 1950. Selected Invertebrate Types. Wiley, New York. 597p.
Petrunkevitch A . 1916. Morphology of Invertebrate Types. MacMillan, New York. 263p.
Ruppert EE, Fox RS, Barnes RB. 2004. Invertebrate Zoology, A functional evolutionary approach, 7 th ed. Brooks Cole Thomson, Belmont CA. 963 pp.
Supplies
Dissecting microscope
Compound microscope
Dissecting pan or culture dish
Isotonic magnesium chloride
Living or preserved clam (Mercenaria or Tapes)
Empty, cleaned shell
Pliers
Slides and coverslips
Carmine-seawater suspension
1 % toluidine blue in seawater
If the process of opening living clams is considered to be too difficult, clams can be easily opened by streaming them a stock pot with about an inch of water for about 10 minutes or until the valves gape. This provides quick access to the soft anatomy but, of course, removes many advantages of using fresh material. Cilia will no longer be beating and textures, colors, sizes and shapes will be altered. Even so, steamed clams are preferable to preserved.