Invertebrate Anatomy OnLine
Littorina irrorata ©
Periwinkle
3jul2006
Copyright 2003 by
Richard Fox
Lander University
Preface
This is one of many exercises available from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises can be accessed by clicking on the links to the left. A glossary and chapters on supplies and laboratory techniques are also available. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Mollusca P, Eumollusca, Conchifera, Ganglionura, Rhacopoda, Gastropoda C, Prosobranchia sC, Caenogastropoda O, Mesogastropoda sO, Littorinoidea SF, Littorinidae F (Fig 12-125)
Mollusca P
Mollusca, the second largest metazoan taxon, consists of Aplacophora, Polyplacophora, Monoplacophora, Gastropoda, Cephalopoda, Bivalvia, and Scaphopoda. The typical mollusc has a calcareous shell, muscular foot, head with mouth and sense organs, and a visceral mass containing most of the gut, the heart, gonads, and kidney. Dorsally the body wall is the mantle and a fold of this body wall forms and encloses that all important molluscan chamber, the mantle cavity. The mantle cavity is filled with water or air and in it are located the gill(s), anus, nephridiopore(s) and gonopore(s). The coelom is reduced to small spaces including the pericardial cavity containing the heart and the gonocoel containing the gonad.
The well-developed hemal system consists of the heart and vessels leading to a spacious hemocoel in which most of the viscera are located. The kidneys are large metanephridia. The central nervous system is cephalized and tetraneurous. There is a tendency to concentrate ganglia in the circumenteric nerve ring from which arise four major longitudinal nerve cords.
Molluscs may be either gonochoric or hermaphroditic. Spiral cleavage produces a veliger larva in many taxa unless it is suppressed in favor of direct development or another larva. Molluscs arose in the sea and most remain there but molluscs have also colonized freshwater and terrestrial habitats.
Eumollusca
Eumollusca is the sister taxon of Aplacophora and includes all molluscs other than aplacophorans. The eumolluscan gut has digestive ceca which are lacking in aplacophorans, the gut is coiled, and a complex radular musculature is present.
Conchifera
Conchifera, the sister taxon of Polyplacophora includes all Recent molluscs other than aplacophorans and chitons. The conchiferan shell consists of an outer proteinaceous periostracum underlain by calcareous layers and is a single piece (although in some it may appear to be divided into two valves). The mantle margins are divided into three folds.
Ganglioneura
Most Recent molluscs are ganglioneurans, only the small taxa Aplacophora, Polyplacophora, and Monoplacophora are excluded. Neuron cell bodies are localized in ganglia.
Rhacopoda
The mantle cavity is posterior in the ancestor although it may be secondarily moved anterior by torsion. This taxon includes gastropods and cephalopods.
Gastropoda C
Gastropoda is the largest molluscan taxon and is the sister group of Cephalopoda. Gastropods are united by descent from a torted ancestor although many exhibit various degrees of detorsion. Many are coiled and asymmetrical but the ancestor was probably symmetrical. Gastropods are relatively unspecialized molluscs known collectively as snails. The univalve shell, present in the ancestral gastropod and in most Recent species, is reduced or lost in many representatives. The flat creeping foot was inherited from their eumolluscan ancestors but gastropods have developed a distinct head with an abundance of sophisticated sense organs. The originally posterior mantle cavity has become anterior as a consequence of torsion, although detorsion has reversed this condition in many. Gastropods were originally gonochoric and most remain so but many derived taxa are hermaphroditic. Most are marine but many taxa have invaded freshwater and the only terrestrial molluscs are gastropods. Most have a single gill, atrium, and nephridium but the most primitive representatives have two of each. Only one gonad, the right, is present. The ancestor probably had an operculum. The nervous system is streptoneurous (twisted by torsion).
Prosobranchia sC
Prosobranchia was once one of three great gastropod subclasses but it is no longer considered to be a monophyletic taxon although the concept continues to be used as a pedagogical convenience. Prosobranchs are the gastropods most like the ancestral snails. They are torted and most have a shell and are coiled and asymmetrical. The mantle cavity is anterior. Most are gonochoric and most have an operculum. Most have only one gill in the mantle cavity but some primitive taxa have two. The right atrium is lost in most. Prosobranchs are specialized for life in marine benthic habitats although representatives are also found in freshwater and on land.
Caenogastropoda O
Caenogastropoda includes the two large and successful groups, mesogastropods and Neogastropoda. One gill, one nephridium, and one atrium are present. The gill is monopectinate, with filaments on only one side of the central axis. This new gill is less prone to fouling with sediment and silt and is probably largely responsible for the success of these snails as it allowed invasion of soft-bottom habitats.
Mesogastropoda sO
Although Mesogastropoda is no longer thought to be monophyletic it remains a useful pedagogic device and the concept is still widely employed. The mesogastropod osphradium is a simple ridge and the radula is taenioglossate, with seven teeth in each row. Mesogastropods occur in marine, freshwater, and terrestrial habitats but most are marine.
Laboratory Specimens
Littorina littorea, the European periwinkle, is typical of mesogastropod snails and is a good example of an unspecialized prosobranch gastropod. This species is readily available, being common on rocky coasts in Europe and the American northeast above the Middle Atlantic States. Littorina is harvested commercially in Europe where it is used for food. Some species of Littorina is available on almost all coasts and living and preserved specimens are sold by supply companies.
The following anatomical account applies specifically to Littorina littorea but can be used with any other member of the genus with little change. Littoraria irrorata (formerly Littorina irrorata), the marsh periwinkle, is common in the Spartina salt marshes of the southeastern United States (photo at http://www.jax.shells.org/litor.htm). Both species achieve sizes of 2-3 cm and are large enough for dissection in introductory invertebrate zoology courses. Other eastern North American species are a little too small for this purpose.
This exercise emphasizes externally visible features and no dissection, other than removal of the shell and opening of the mantle cavity, is required. A supplementary optional study of internal anatomy is included.
If possible study a living snail that has been relaxed by the gradual addition of concentrated magnesium chloride (see below). A preserved snail may also be used. Relaxed animals should be observed in a small pan or dish of isotonic magnesium chloride (see Supplies and Recipes chapter) whereas preserved animals should be in tapwater. The study should be conducted with a dissecting microscope and the animal should be in a small dissecting pan. Color descriptions apply to living specimens.
Anesthetization
Shelled molluscs are notoriously difficult to anesthetize. Upon detecting anesthetic in their water they immediately withdraw, close the shell, and isolate themselves from the anesthetic. Relaxation is important, however, as it is very difficult to remove imperfectly relaxed animals from their shells and difficult to dissect them after removal. Special gradual relaxation techniques must be employed.
Snails should be relaxed by the teaching staff prior to the laboratory meeting. The snails should be placed in a small container of seawater about 24 hours prior to the start of the laboratory period. There should be just enough seawater to cover the animals. Concentrated magnesium chloride (70 parts per thousand) should be added dropwise to the seawater containing the animals. Arrange a slow siphon or drip-string to drip 70 ‰ magnesium chloride slowly into the container. Avoid disturbing the animals until it is time to use them.
Externally Visible Features
Shell
Look at the shell first (Fig 1, 12-27A,B). It is heavy and smooth, without sculpture or ornamentation. It is coiled in a series of complete turns, or whorls, of increasing diameter around a central axis known as the columella (Fig 12-27A). The columella is not visible externally. The last and by far the largest whorl is the body whorl. Most of the soft parts of the snail are contained in the body whorl. The combined smaller whorls sitting atop the body whorl together make up the spire.
Figure 1. The shell of the periwinkle, Littorina littorea. Gastrop154L.gif
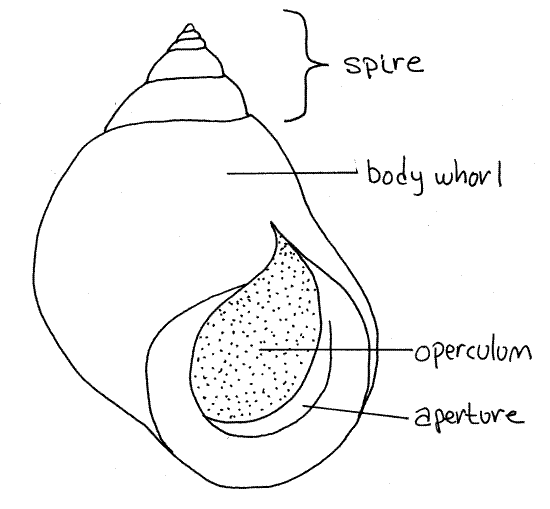
The body whorl terminates at a large oval opening, the aperture from which the foot and head of the animal can be extended (Fig 1, 12-27A,B). In Littorina, and most other mesogastropods, the aperture is on the right of the columella when facing the observer if the spire is pointed up. Such an arrangement is said to be dextral, or right handed (Fig 1). The opposite is sinistral, in which the aperture is on the left of the columella. The rim of the aperture is smooth.
Soft Anatomy
Find the thin, disc-like, proteinaceous operculum that serves as a door to close the aperture when the soft parts are withdrawn into the shell (Fig 2, 12-27C-F). It is attached to the thick muscular foot (Fig 2, 12-27C-F). The foot is attached to the columella by the powerful columellar muscle which cannot be seen at present. With forceps pull gently on the operculum to expose as much of the animal as possible.
In life, the pale ventral sole of the foot makes contact with the substratum and is used for crawling. In your specimen it may be folded on itself as it always is when retracted (Fig 3). The sole of the foot is the ventral surface of the animal.
Figure 2. The left side of a male Littorina littorea from Pemaquid Point, Maine. The shell has been removed but the mantle is intact. Structures in the mantle cavity are viewed through the mantle. Gastrop155La.gif
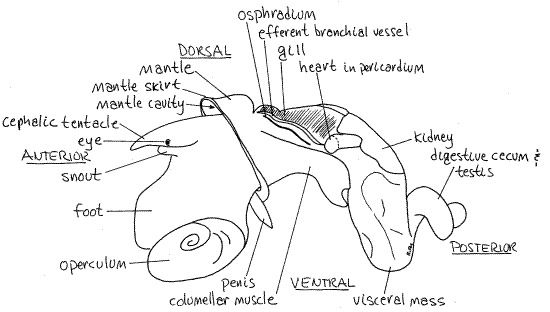
The head is an anterior protuberance dorsal to the foot (Fig 2). It bears two sensory cephalic tentacles with a lateral eye on the base of each. A median snout protrudes anteriorly from between the bases of the two tentacles (Fig 2). The mouth is located at the anterior end of the snout. The snout is not retractile and is thus not a proboscis.
Posterior to the head, on the dorsal side of the animal, is the mantle cavity (Fig 2). It is a deep pocket above and behind the head. The mantle cavity is enclosed by the mantle, whose anterior edge is the mantle skirt. The skirt forms a collar (Fig 3) that completely encircles the body behind the head and lies against the lips of the aperture of the shell. You may be able to see the delicate gill, on the left side of the mantle cavity (Fig 2) and, if the specimen is a male, a penis on the right (Fig 2).
Use the features you have found so far to orient the animal. The foot is ventral, the head anterior, and the mantle cavity dorsal and posterior to the head. The snail is coiled and asymmetrical. The remainder of the body is the visceral mass but it is coiled up into the shell and cannot be seen at present.
" To remove the animal from the shell it is necessary that the columellar muscle be detached from the columella. This muscle extends from the columella to the foot and is the muscle used by the animal to retract its foot and head into the shell. It is also the connection that holds the animal firmly in its shell and as such it must be detached from the columella. This muscle, however, is not yet accessible and part of the shell must be removed before the muscle can be detached.
Littorina and other small or thin-shelled species are best extracted from their shells by cracking the shell with a small C-clamp or vise. The shell is removed carefully and in pieces from around the animal until the columellar muscle can be separated from the columella. The snail can then be removed easily from the remains of the shell.
Place the snail in a 4-inch C-clamp or small vise and apply pressure until the shell cracks. You can hear and feel it crack. Remove the shell from the clamp and see if any pieces around the aperture can be removed without damage to the soft anatomy. If so, lift them with forceps and discard them. Put the shell back in the clamp and apply pressure to an as yet uncrackedportion of the shell until it cracks. Never close the clamp on soft parts or apply additional pressure after the shell cracks. Pressure should be applied only to sound, uncracked parts of the shell. Remove any loose pieces of shell as before.
Continue this until you can see the white columellar muscle extending from the foot to the columella of the shell (Fig 3). Use a scalpel to scrape (rather than cut) the columellar muscle from the shell. Be sure to break all connections between the muscle and the columella.
It should now be possible to unwind the snail from the remains of the shell. Do not force the snail out if the shell if it does not move easily. The soft tissues of the visceral mass tear easily but if that happens, the subsequent dissection will be more difficult. If the snail does not come easily out of the shell, check to be sure you have completely detached the columellar muscle. If not, do so. It may be necessary to continue cracking the shell to remove more shell until the snail can be extracted.
When you have removed the snail from its shell, review the anatomical features you found earlier (Fig 2, 3). Find the head, foot, sole, tentacles, eye, mouth, operculum, mantle, skirt, and mantle cavity again. Be sure you can still orient the animal.
Quickly survey those parts of the snail that until now were concealed by the shell. The visceral mass was formerly coiled up into the shell. It is now seen as a soft and delicate coiled mass that must be handled carefully. Its walls are thin and transparent and you can see many of its structures through them without dissection.
Mantle Cavity
The mantle, which is the dorsal body wall, forms a fold that is the roof and walls of the mantle cavity. The mantle cavity is a deep pocket lying dorsally, above the anterior part of the visceral mass, posterior to the head. Its only opening lies above the head and is bordered by the mantle skirt (Fig 3). The anterior edge of this mantle fold is the skirt. Before extraction, the skirt lined the lip of the aperture. Gently slip a blunt probe into the opening to see how deep the cavity is.
Look through the transparent roof of the mantle cavity and find the most conspicuous structures inside it before you open it. Starting on the left is the osphradium (Fig 2, 4). In Littorinait is simply a pigmented ridge of sensory epithelium. It is not an elaborate gill-like structure as it is in neogastropods such as Busycon and Ilyanassa.
The gill is a large, long, pale brown, oval organ lying medial to the osphradium. It extends from the osphradium to the midline and from the anterior to the posterior ends of the mantle cavity. Its left border bears faint white pigmentation that marks the location of the efferent branchial vessel which drains oxygenated blood from the gill to the heart. Only the left gill is present in caenogastropods and it is monopectinate.
The hypobranchial gland, which secretes mucus into the mantle cavity, is a long narrow secretory region of the mantle roof located on the right side of the gill. It is about the same size as the gill. Other features of the mantle cavity will be observed after it is opened.
The intestine exits the stomach and winds through the visceral mass where it is hidden from view. Its posterior end, the rectum, emerges from the mass and extends obliquely across the roof of the mantle cavity on the right border of the hypobranchial gland (Fig 3, 4). It is easily seen in males, but is hidden from view in reproductive females by the large, yellowish egg capsule gland on the right edge of the mantle cavity. The prostate gland, which occupies the equivalent position in the male, is not as large and does not hide the rectum.
Figure 3. The right side of a male Littorina littorea with the shell removed. Gastrop156La.gif
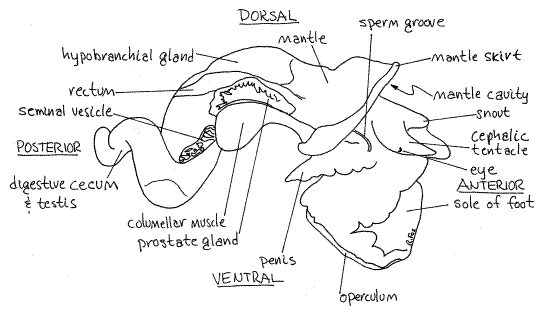
The large, white, right columellar muscle, which you separated from the columella in order to remove the shell, is on the right side of the body (Fig 3). It extends from the columella to the foot. When it contracts, the animal is withdrawn into the shell. The columellar muscle is homologous to a pedal retractor muscle of ancestral molluscs and only the right one is present. There is no left columellar muscle. The distal part of the reproductive and digestive systems lie between the hypobranchial gland and the columellar muscle.
The kidney, or nephridium, is the large, pinkish-brown organ lying on top of the visceral mass immediately posterior to the mantle cavity (Fig 2). To the left of it is the pericardial cavityand heart. The heart and kidney are immediately posterior to the posterior end of the gill.
The remainder of the visceral mass is occupied chiefly by the dark brown digestive cecum but the gonad is mixed with it on the right side and the stomach and intestine are surrounded by it. Part of the black stomach can be seen on the left side of the visceral mass posterior to the heart and kidney.
" Open the mantle cavity with a dorsal, median, longitudinal incision (with fine scissors) through its roof. Begin at the anterior margin of the mantle skirt and cut all the way to the posterior end of the cavity. Cut between the gill and the hypobranchial gland. The cut will not be precisely median rather will run obliquely across the roof. Deflect the two sides of the roof and look inside. Find once more the osphradium, gill, and hypobranchial gland (Fig 4). If you have a male, the large, black penis will be visible lateral to the right tentacle.
The gill is monopectinate, with filaments on only one side of its long axis. The efferent branchial vessel is contained in its axis. On the right side of the mantle cavity find the large, tubular, transparent rectum running obliquely across the roof beside the hypobranchial gland. The rectum narrows near its end to form an anal papilla that projects into the right mantle cavity. The anus is located at the free end of the papilla. The anus opens into the right side of the mantle cavity.
If you have a male, you will see the large, yellowish prostate gland on the right of the rectum (Fig 3). If your animal is a female there will be an egg capsule gland in about the same position beside the rectum (Fig 4). The female gonopore opens at the anterior tip of the egg capsule gland. The gonopore empties into the right side of the mantle cavity.
The large pink structure visible on the floor of the mantle cavity on the left is the esophageal gland. This is an expansion of the walls of the esophagus.
Excretory System
Relocate the pinkish-brown kidney, on the dorsal surface of the visceral mass adjacent to the posterior end of the mantle cavity (Fig 4). Look at its anterior surface from inside the mantle cavity and find the large, slit-shaped nephridiopore. An ultrafiltrate of the blood is produced by ultrafiltration of the blood into the pericardial cavity. A nephrostome in the wall of the pericardial cavity opens into the renopericardial canal which leads into the kidney. Blood in the surrounding hemocoel bathes the outside of the kidney and canal and modifies the ultrafiltrate inside by selective absorption and secretion. Final urine is released into the back of the mantle cavity from the nephridiopore.
Hemal System
Find the pericardial cavity again (Fig 2, 4). It is a small coelomic compartment immediately posterior to the posterior end of the gill and on the left edge of the kidney. Notice that the efferent branchial vessel from the gill runs into its anterior corner.
The heart is the brown organ within the pericardial cavity. It is composed of one atrium and a ventricle. The ventricle is posterior to and larger than the atrium. The atrium is small, thinner walled, and anterior. It is a little darker than the ventricle.
Oxygenated blood from the gill enters the atrium via the efferent vessel and then flows into the lumen of the ventricle (Fig 12-52). The large, transparent, inconspicuous aorta exits the posterior end of the ventricle. It divides immediately into anterior and posterior aortae.
Contractions of the muscular walls of the ventricle force blood into the aortae and thence to the hemocoelic spaces. You may wish to open the pericardial cavity by removing the thin, membranous pericardium from the heart. This will make it easier to distinguish clearly between the atrium and ventricle.
Nervous System
The nervous system cannot be observed without further dissection (Fig 12-53). If study of this system is desired the few comments included in the optional section on internal anatomy may prove useful. For a more thorough treatment of the nervous system the student should refer to Fretter and Graham (1994).
Figure 4. Dorsal view of a female Littorina littorea with the mantle cavity and hemocoel opened. The mantle cavity has been opened by an incision between the gill and hypobranchial gland. The left side of the mantle roof has been deflected to the left and the right side to the right. The hemocoel has been opened by a mid-dorsal incision through the body wall under the mantle cavity. Gastrop157La.gif
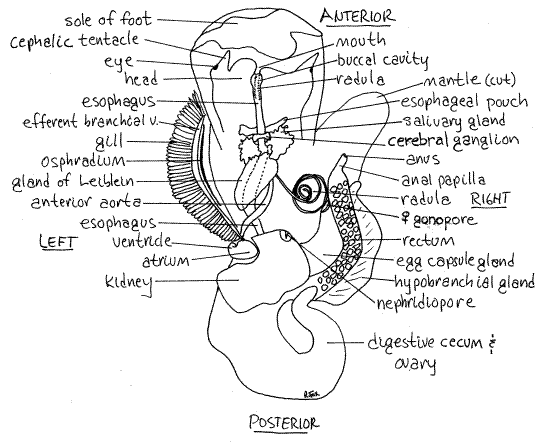
Reproductive System
During the breeding season the reproductive system is fully developed and easily studied but at other times the genital organs diminish in size and are difficult to find. The sexes are separate (gonochoric), and fertilization is internal. As usual in gastropods with internal fertilization, the gonoducts are complicated and regionally specialized. There is a pelagic veliger larva.
Most of the reproductive system of most prosobranchs can usually be seen from the exterior and dissection is not required to study its major features. This is the case with Littorina.
The reproductive organs are located on the right side of the body and in both sexes consists of a gonad (ovary or testis) located on the right, or concave, side of the upper coils of the visceral mass and an elaborate duct connecting the gonad with the gonopore in the mantle cavity. The gonad occupies a coelomic space, the gonocoel.
Fertilization is internal with copulation and the gonoduct is complicated. It is composed of three sections derived, in order from proximal to distal, from the old coelomoduct of the gonocoel, part of the right nephridium and its duct, and modified areas of the mantle epithelium.
Male
The most obvious feature of the male reproductive system is the large, black penis on the right side of the head (Fig 3). Look for it lateral and posterior to the right cephalic tentacle and use its presence or absence to determine the sex of your specimen.
If you have a male, look at the posterior end of the visceral mass to find the testis (Fig 3, 12-56B). Its gray or gray-green lobes are interspersed between the brown lobes of the digestive ceca and the two may be difficult to distinguish. A conspicuous (in season), convoluted seminal vesicle runs along the right side of the visceral mass then along the right edge of the mantle cavity to enter the prostate gland (Fig 3, 12-56B). This large, cream or yellow gland lies on the right edge of the rectum.
Look inside the mantle cavity for the raised sperm duct running longitudinally along the center of the surface of the prostate gland. This duct connects with a ciliated sperm groove that extends across the floor of the mantle cavity to the base of the penis (Fig 3, 12-56B).
Female
The ovary is pink, rose, or yellowish and its small lobes are mingled with those of the digestive gland in the visceral mass (Fig 4, 12-56A). The narrow oviduct is pink when it contains eggs and lies low on the right side of the visceral mass. It runs into a cream-yellow complex of reproductive organs dorsal to the columellar muscle. The posterior part of this complex is thealbumen gland. The larger anterior part is the egg capsule gland (Fig 4). The distal oviduct exits the anterior end of the egg capsule gland and opens via the female gonopore on the right side of the mantle cavity (Fig 4, 12-56A).
Your study of Littorina may end here if desired.
Internal Anatomy
If you wish to continue, the best way to gain a general understanding of internal anatomy is by tracing the gut posteriorly from the mouth through the visceral mass to the anus.
" Insert one point of your fine scissors into the mouth and cut posteriorly along the dorsal midline of the head. When you reach the mantle cavity remove the scissors from the gut but continue cutting, now through the floor of the mantle cavity to expose the large hemocoel located below it. Stop cutting when you reach the posterior end of the mantle cavity. Deflect the cut edges of the floor of the mantle cavity and pin them out of the way if necessary.
Digestive System
Return to the head and study the anterior end of the gut tube. Part of it has been opened and its lumen exposed by your incision. The mouth at the tip of the snout opens into a shortbuccal cavity which soon becomes the long tubular esophagus (Fig 4, 12-53). The region between the buccal cavity and esophagus is sometimes called the pharynx.
Find the toothy ribbon of the radula in the radular sac ventral to the buccal cavity and esophagus (Fig 4, 12-53). The sac and the radula it contains are exceedingly long in this species and extend far posteriorly into the hemocoel, which you just opened.
The thin-walled esophagus lies above the radula in the head and anterior visceral mass but it may be difficult to see because of its transparency (Fig 4). Trace the esophagus posteriorly from the head into the hemocoel. Two pairs of glands empty into it.
Anteriormost are the two small esophageal pouches of unknown function (Fig 4). They are pale, whitish, and lie beside the esophagus just anterior to the hemocoel.
Posterior to them are the two much larger, frilly, bright yellow salivary glands (Fig 4). They empty via long ducts into the gut anterior to the esophageal pouches. Consequently, as evaginations of the gut, the salivary glands are actually anterior to the esophageal glands but because of their long ducts, they can occupy a more posterior position.
The cerebral commissure between the two cerebral ganglia crosses the esophagus dorsally between the esophageal pouches and the salivary glands (Fig 4).
The esophagus posterior to the salivary glands has walls thickened with purplish secretory tissue referred to as the esophageal gland or gland of Leiblein (Fig 4).
The anterior aorta is a large white tube on the right of the esophagus (Fig 4). Do not confuse it with the esophagus. The coiled radula and radular sac also lie to the right of the esophagus.
The esophagus turns to the left of the midline and exits the posterior left corner of the hemocoel to enter the visceral mass on its left side behind the heart.
" From this point on trace the gut by opening it with your fine scissors. Insert one point of the scissors into the gut lumen and cut through the intervening layers of tissue to do this. In this way you can follow its lumen through the visceral mass.
The esophagus enters the visceral mass as a black tube partly exposed on the surface of the mass. It soon empties into the elongate stomach. Because torsion has bent the gut into a U, the esophagus enters the middle of the stomach rather than its anterior end. Part of the stomach is visible on the surface of the left side of the anterior visceral mass.
" Open the stomach. The ducts of the digestive ceca can be seen entering the stomach near the point where the esophagus enters. The anterior end of the stomach is the style sac whose ciliated walls rotate the stomach contents, which is a soft rod known as the protostyle. The lumen of the stomach is divided into two chambers by a longitudinal fold of tissue arising beside the opening of the esophagus and passing posteriorly. This fold forms a chamber, sometimes called the gastric cecum, which receives food directly from the esophagus. The food moves from gastric cecum into the stomach proper where it is exposed to enzymes from the digestive ceca, is mixed by muscular contractions of the stomach walls, and rotated by cilia.
Note the two typhlosoles, or folds, on the stomach wall. The ciliated intestinal groove lies between the two typhlosoles. The typhlosoles and intestinal groove arise in the style sac of the stomach and extend into the intestine. The intestinal groove originates at a sorting field of ciliated ridges and grooves located beside the openings of the digestive glands. The intestineexits the anterior end of the stomach.
" Trace the intestine by cutting through the viscera and opening its lumen.
The intestine turns posteriorly into the interior of the visceral mass, then turns again and runs anteriorly as the rectum. The rectum crosses the roof of the mantle cavity where you have already seen it. If the intestine contains fecal pellets it is easily traced.
Nervous System
The nerve ring consisting of several pairs of fused ganglia lies around the esophagus between the esophageal pouches and salivary glands. The two large cerebral ganglia lie beside the esophagus and are connected with each other by the cerebral commissure across the top of the esophagus (Fig 4, 12-53). Other ganglia, commissures, and connectives complete the nerve ring around the ventral side of the esophagus.
Radula
>1a. Unwind the radula to its full extent and compare its length with the length of the snail. <
>1b. Remove the radula and make a wetmount for examination of the radula with the compound microscope. Mesogastropods have taenioglossate radulae in which the teeth are in transverse rows of seven teeth each (Fig 12-40C). Each row consists of a central median tooth with three lateral teeth on each side. <
References
Abbott DP . 1987. Observing Marine Invertebrates. Stanford Univ. Press, Stanford. 380p. Littorina planaxis.
Fretter V, Graham A. 1994. British Prosobranch Molluscs, 2 nd ed. Ray Society, London. 820 pp. (With an extensive treatment of Littorina littorea, pp 14-48)
Ruppert EE, Fox RS, Barnes RB. 2004. Invertebrate Zoology, A functional evolutionary approach, 7 th ed. Brooks Cole Thomson, Belmont CA. 963 pp.
Supplies
Dissecting microscope
Dissecting set with microdissecting tools
living or preserved Littorina
small dissecting pan
4" C-clamp
Isotonic magnesium chloride
#1 insect pins