Invertebrate Anatomy OnLine
Ctenocephalides ©
Cat and dog fleas
30jun2006
Copyright 2001 by
Richard Fox
Lander University
Preface
This is an exercise from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises can be accessed by clicking on the links to the left. A glossary and chapters on supplies and laboratory techniques are also available. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Arthropoda P, Mandibulata sP, Tracheata, Hexapoda SC, Insecta C, Dicondylia, Pterygota, Metapterygota, Neoptera, Eumetabola, Holometabola, Siphonaptera O, Pulicoidea SF, Pulicidae F, Archaeopsyllinae sF, (Fig 16-15, 20-14, 20-15, 21-23)
Arthropoda P
Arthropoda, by far the largest and most diverse animal taxon, includes chelicerates, insects, myriapods, and crustaceans as well as many extinct taxa. The body is segmented and primitively bears a pair of jointed appendages on each segment. The epidermis secretes a complex cuticular exoskeleton which must be molted to permit increase in size. Extant arthropods exhibit regional specialization in the structure and function of segments and appendages. The body is typically divided into a head and trunk, of which the trunk is often itself divided into thorax and abdomen.
The gut consists of foregut, midgut, and hindgut and extends the length of the body from anterior mouth to posterior anus. Foregut and hindgut are epidermal invaginations, being derived from the embryonic stomodeum and proctodeum respectively, and are lined by cuticle, as are all epidermal surfaces. The midgut is endodermal and is responsible for most enzyme secretion, hydrolysis, and absorption.
The coelom is reduced to small spaces associated with the gonads and kidney. The functional body cavity is a spacious hemocoel divided by a horizontal diaphragm into a dorsal pericardial sinus and a much larger perivisceral sinus. Sometimes there is a small ventral perineural sinus surrounding the ventral nerve cord.
The hemal system includes a dorsal, contractile, tubular, ostiate heart that pumps blood to and from the hemocoel. Excretory organs vary with taxon and include Malpighian tubules, saccate nephridia, and nephrocytes. Respiratory organs also vary with taxon and include many types of gills, book lungs, and tracheae.
The nervous system consists of a dorsal, anterior brain of two or three pairs of ganglia, circumenteric connectives, and a paired ventral nerve cord with segmental ganglia and segmental peripheral nerves. Various degrees of condensation and cephalization are found in different taxa.
Development is derived with centrolecithal eggs and superficial cleavage. There is frequently a larva although development is direct in many. Juveniles pass through a series of instars separated by molts until reaching the adult size and reproductive condition. At this time molting and growth may cease or continue, depending on taxon.
Mandibulata sP
Mandibulata includes arthropods in which the third head segment bears a pair of mandibles. As currently conceived this taxon includes myriapods, hexapods, and crustaceans. Appendages may be uni- or biramous and habitats include marine, freshwater, terrestrial, and aerial.
Tracheata
Myriapods and hexapods share tracheae and a single pair of antennae and are sister taxa in Tracheata. Crustaceans, which have gills and lack tracheae, are excluded and form the sister group.
Hexapoda SC
The body is divided into three tagmata; head, thorax, and abdomen. Appendages are uniramous and a single pair of antennae is present. Three pairs of legs and two pairs of wings are found on the thorax of most adults. Hexapod legs are uniramous although there is increasing evidence that they evolved from multiramous appendages of their ancestors. Gas exchange is accomplished by trachea. Excretory organs are Malpighian tubules and the end product of nitrogen metabolism is uric acid. There is relatively little cephalization of the nervous system. Insects are gonochoric with copulation and internal fertilization.
Insecta C
Most hexapods are insects. A few hexapod taxa (orders) lack wings and have primitive mouthparts recessed into the head and belong to Entognatha, the sister taxon of Insecta. Insects have ectognath mouthparts and the adults (imagoes) of most taxa have wings.
Pterygota
The winged insects. These insects are derived from a winged common ancestor. Adults of most taxa have wings although they have been lost in some.
Eumetabola
Juveniles have no ocelli and there are six or fewer Malpighian tubules.
Holometabola
The final larval instar pupates and undergoes a radical metamorphosis in which it is converted to an imago, or adult. The imago is sexually mature and in most taxa has wings whereas larvae are immature and wingless. During metamorphosis many or most larval tissues are dismantled and adult structures built anew. Wings, for example, are manufactured from clusters of undifferentiated cells known as imaginal discs but not from preformed wingpads as in pauro- and hemimetabolous insects.
Siphonaptera O
Fleas are small, secondarily wingless, laterally compressed blood sucking insects. Most fleas are mammalian ectoparasites but about 5% parasitize birds. About 2400 extant species are known. Like most parasites, fleas are highly derived and specialized. The mouthparts are adapted for piercing and sucking and mandibles are lacking. Because of their highly derived condition zoologists have encountered difficulties in recognizing homologies between flea mouthparts and those of other insects. A blood meal is required prior to oviposition. Many are disease vectors. The cuticle is sclerotized and hard and the entire body surface is protected by sclerites. A pair of ocelli may be present but compound eyes are absent. Development is holometabolous and the larvae are slender and wormlike. Larvae are not parasitic and feed on organic detritus, including partially digested host blood in the feces of the adults. Fleas are adapted for passing rapidly and unimpeded through the hair or feathers of the host. Adaptations for this are the smooth, compressed body, absence of wings, posteriorly pointing spines and setae, and short antennae that can be sequestered in a fossa on the side of the head. Attempts by the host to remove the flea are impeded by the backward pointing setae and spines. Fleas arose from winged ancestors and are probably related to Mecoptera (Scorpion flies) and Diptera (flies).
Laboratory Specimens
Cat fleas, Ctenocephalides felis, and dog fleas, Ctenocephalides canis, can be collected from a cooperative pet in quantities limited only by the pet’s (and collector’s) patience. The two species are similar, in fact are difficult to distinguish from each other, and either is suitable for this exercise. Both are cosmopolitan and both occur on cats and dogs, and occasionally humans. The human flea, Pulex irritans, is also cosmopolitan and belongs to the same family (Pulicidae). Cleared wholemount slides of C. felis, C. canis, P. irritans, and Xenopsylla cheopis(the rat flea and also a member of the same family) are available from biological supply companies. Unmounted or mounted specimens can be used for this exercise. Unmounted specimens should be observed with the dissecting microscope, wholemounts with the compound microscope. The dog flea, C. canis, is the intermediate host of the dog tapeworm, Dipylidium caninum, found in both dogs and cats. The rat flea, X. cheops, is a vector of bubonic plague. Haplopsyllus anomalus, another pulicid, is a parasite of California ground squirrels and is also a plague vector. In the eastern United States Ctenocephalides felis is the species most often found in human dwellings whereas on the Pacific coast it is Pulex irritans.
External Anatomy
Tagmata
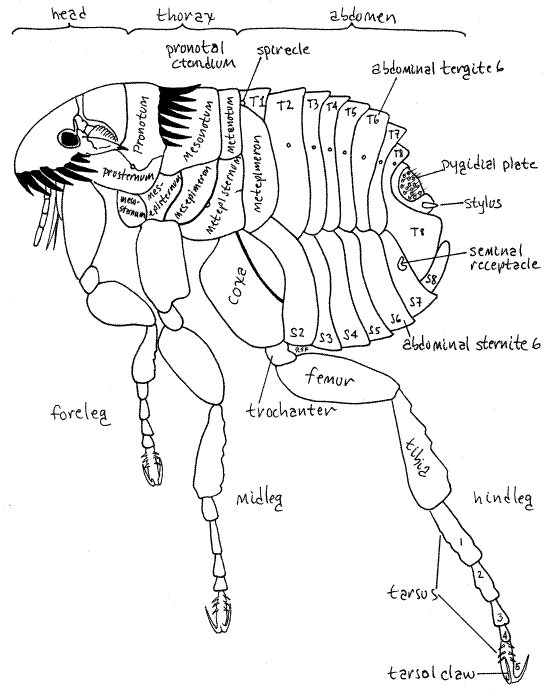
Like other insects, fleas have a body consisting of three tagmata; head, thorax, and abdomen (Fig 1, 21-22A). Unlike most other insects, however, the distinction between the tagmata is not distinct and it is not obvious at a glance where one tagma bends and another begins.
Figure 1. The cat flea, Ctenocephalides felis. Setae have been omitted for clarity. Siphon13L.gif
Head
Head Capsule
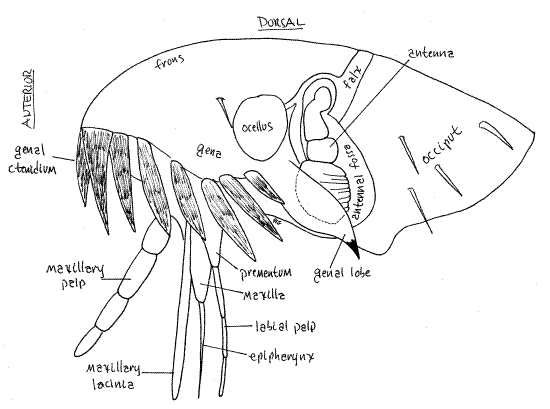
Like that of other insects, the head is enclosed in a sclerotized cranium known as the head capsule (Fig 2). The capsule is a single unit formed by the fusion of the sclerites of the head segments, which are themselves no longer recognizable. The head capsule is divided into anterior and posterior regions by a thick cuticularized ridge, the falx, arching transversely from one side of the head to the other.
The head capsule consists of the typical regions characteristic of insects. The front of the head, anterior to the falx, is the frons. The top of the head is the vertex. The posterior region, near the thorax, is the occiput. The gena is ventral and posterior to the ocellus.
The articulation between the anterior head capsule and the posterior region is slightly flexible in some taxa (Fracticipita sO) but not in others (Integricipita sO) such as Ctenocephalidesand other pulicids. The falx marks the zone of articulation between the two regions.
In Ctenocephalides the ventral margin of the gena bears agenal ctenidium (ctene = comb) consisting of a comblike row of strong, stout, heavily sclerotized spines (Fig 1,2). OnlyCtenocephalides have genal ctenidia with more than five spines (Fig 1).
Figure 2. Lateral view of the head of a cat flea, Ctenocephalides felis. Siphon14L.gif
Eyes
Compound eyes are lacking. Lateral ocelli present in some taxa, including Ctenocephalides (Fig 2). The ocellus has a single lens.
Antennae
Ventral to the falx is a lateral depression, the antennal fossa, in which the antenna are sequestered. The antenna can be moved out of the way into the fossa so they do not get snagged on the hair of the host. A strong, spine-tipped genal lobe projects posteriorly from the gena over the antennal fossa to further protect the antenna from snagging on the pelage of the host.
The antennae are multiarticulate but the terminal nine or so articles are short, wide, and appear to form a single large, clublike article, known, appropriately, as the club Fig 2). The two proximal articles are large and distinct. The antenna is in an unusual position on the side of the head where it lies out of the way in a recess, the antennal fossa. During copulation males deploy their antennae and use them to hold the female.
Mouthparts
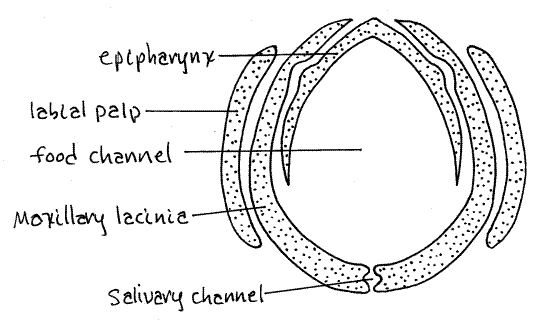
Siphonapteran mouthparts are adapted for piercing and sucking (Fig 2, 3). The long slender stylet, which is the piercing organ, is formed of two slender, bladelike maxillary lacinia and a needlelike epipharynx (Fig 2). The stylet is supported by the labium. The labrum and hypopharynx are vestigial and mandibles are absent.
The unpaired, median epipharynx extends ventrally from the lower surface of the head, anterior to the mouth (Fig 2,3). In the past it was thought to be the labrum but it is now considered to be an outgrowth of the head capsule. It is long slender and bladelike.
The maxilla bears two laciniae and two maxillary palps with 4-5 articles (Fig 2). The maxillary laciniae are long and slender and form the outer part of the stylet (Fig 3). The serrated, sawlike laciniae are used to cut into the skin of the host, permitting the epipharynx to slip into a blood vessel. In the past the laciniae were thought to be the highly modified mandibles but it is now believed that mandibles are absent in fleas. Linnaeus believed the maxillary palp, because of its morphology and position, to be the antenna.
The two laciniae and the epipharynx together enclose a food channel for inbound blood (Fig 3). The laciniae form a smaller salivary channel for outbound saliva. The maxillae have antenniform, sensory maxillary palps.
The labium is represented by a proximal prementum and two 3-4 articulate labial palps which form a sheath around the stylet (Fig 2,3). The labium is not part of the stylet and it does not enter the host. Instead, it supports and guides the stylet.
Suction to pull blood into the stylet and gut is provided by cibarial and pharyngeal pumps. The walls of the cibarium (Fig 21-7) and of the pharynx have radial muscles whose contractions dilate their respective lumina to create suction.
Thorax
The three thoracic segments (prothorax, mesothorax, and metathorax) are subequal in size but are not as easily recognized as are those of other insects. Each bears a pair of legs and is covered by several sclerites. Dorsally the prothorax, mesothorax, and metathorax are covered by the pronotum, mesonotum, and metanotum, respectively. These three sclerites are the best landmarks for recognition of the segmentation of the thorax.
Figure 3. Diagrammatic representation of a generalized flea stylet in cross section. Siphon15L.gif
Ventrally the segments are protected by sternites. The large prosternum extends under the head capsule, giving the impression that the forelegs arise from the head (Fig 1). Ctenocephalides has a conspicuous pronotal ctenidium (similar to the genal ctenidium) along the posterior border of the pronotum (Fig 1,2). Although wings are absent in fleas, some entomologists believe two posterior sclerites, the mesepimeron and metepimeron of the mesothorax and metathorax, respectively, are vestiges of the forewings and hindwings of the winged ancestors.
Legs
Each thoracic segment bears a pair of legs, which resemble each other except that the hindlegs are larger and adapted for jumping (Fig 1). The segments and their legs are; prothorax and forelegs, mesothorax and midlegs, and metathorax and hindlegs. The legs consist of coxa, trochanter, femur, tibia, tarsus and tarsal claws, in sequence from proximal to distal. The coxae and femora are large and flattened. The tarsi comprise five tarsomeres, with large, terminal tarsal claws. The legs are heavily spinose.
Wings
Although fleas are wingless they posses lateral sclerites (discussed above) on the mesothorax and metathorax that may be wing vestiges.
Abdomen
The abdomen is ten-segmented (Fig 1). Most segments are covered by a dorsal tergite and a ventral sternite. Segments 8-10 are the genital segments and the tenth tergite has a complex sclerite, the pygidial plate (= sensillium), equipped with slender trichobothria-like setae apparently sensitive to weak air currents (Fig 1). Ten pairs of stigmata are present, two thoracic and eight abdominal, and open into the tracheal system.
Life Cycle
Adult fleas live on mammals or birds, on whose blood they feed. Mating occurs on the host and eggs are produced to drop from the host into the host’s nest or domicile. First instar larvae hatch from the eggs and feed on organic debris in the nest. The larvae are not parasites and do not live on the body of host, rather live in its nest. The feces of adult fleas contains abundant partially digested host blood which contributes to the supply of organic detritus in the nest and is an important food source for the larvae. Larvae are wormlike, with head capsule, but no legs. Larvae have primitive mandibulate (biting and chewing) mouthparts, unlike those of the adults, and unmodified for piercing and sucking. Eventually the larvae spin a cocoon and pupate, still in the nest. Development is holometabolous and imagoes emerge from the pupa and move onto the host.
References
Anon. 1967. Pictorial keys to arthropods, reptiles, birds, and mammals of public health significance. U.S. Dept. Health, Education, Welfare, Public Health Service, National Communicable Disease Center. 192 pp.
Borrer DJ, Triplehorn CA, Johnson NF. 1989. An introduction to the study of insects, 6 th ed. Saunders, Philadelphia. 875pp.
Chapman RF. 1998. The insects, Structure and function, 4 th ed. Cambridge Univ. Press, Cambridge. 769 pp.
Comstock JH. 1930. An introduction to entomology. Comstock Publishing, Ithaca NY. 1044pp.
Daly HV, Doyen JT, Erhlich PR. 1978. Introduction to insect biology and diversity. McGraw-Hill, New York. 564pp.
Evans HE. 1985. The flea, pp37-47 in: The pleasures of entomology. Smithsonian Inst. Press, Washington. 238 pp.
Furman DP, Catts EP. 1970. Manual of medical entomology, 3 rd ed. National Press Book, Palo Alto, CA. 163 pp.
Ruppert EE, Fox RS, Barnes RB. 2004. Invertebrate Zoology, A functional evolutionary approach, 7 th ed. Brooks Cole Thomson, Belmont CA. 963 pp.
Schmidt GD, Roberts LS. 1989. Foundations of parasitology, 4 th ed. Times Mirror Mosby, St. Louis. 750pp.
Supplies
Compound microscope
Cat or dog flea