Invertebrate Anatomy OnLine
Clypeaster rosaceus ©
Sea Biscuit
25may2007
Copyright 2001 by
Richard Fox
Lander University
Preface
This is one of many exercises available from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises, a glossary, and chapters on supplies and laboratory techniques are also available at this site. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Echinodermata P, Eleutherozoa, Cryptosyringida, Echinozoa, Echinoidea C, Euechinoida sC, Clypeasteroida O, Clypeasteridae F (Fig 9-26, 27-12, 28-62)
Echinodermata P
Echinoderms are secondarily radially symmetric deuterostomes whose ancestors were bilaterally symmetric. The adult radial symmetry is pentamerous with body parts occurring in fives or multiples thereof. Echinoderms have strong affinities with the ancestral trimeric deuterostomes especially in the tripartite organization of the coelomic cavities. Echinoderm larvae have the coelom divided into three regions, as is typical of the early coelomates, and these regions have important adult derivatives. All echinoderms are marine and benthic. About 6000 Recent species are known but the fossil record includes 13,000 extinct species.
An important echinoderm apomorphy is the water vascular system that in most groups functions in support of locomotory tube feet but is also important in gas exchange, excretion, and feeding. The body wall includes a thick connective tissue dermis in which calcareous ossicles (little bones) are almost always an important component. These ossicles make up an endoskeleton which assumes different forms in different taxa. In most echinoderms calcareous spines of various sizes and shapes arise from the dermis and extend from the body surface and are alluded to by the name echinoderm (= spiny skin). The connective tissue is mutable and its consistency is under nervous control.
Excretion in echinoderms is accomplished by simple diffusion of metabolic wastes (ammonia) across thin permeable regions of the body wall. A variety of gas exchange structures, including the tube feet, is found in various echinoderms. A hemal system is present but its role in transport is still poorly understood and the chief transport system is the circulating fluid of the various coelomic compartments. The hemal system may be through transport system that delivers nutrients from the gut to these compartments for local distribution. The nervous system consists of two central intraepidermal nerve rings from which arise radial nerves to the periphery. Echinoderms are gonochoric and fertilization is usually external.
Eleutherozoa
Eleutherozoans are mobile echinoderms in which the oral surface is oriented against the substratum. A madreporite and locomotory tube feet are present. Polian vesicles and Tiedemann’s bodies may be present on the ring canal. Movable spines are present. Eleutherozoa includes all Recent echinoderms except for its sister taxon, Crinoidea.
Cryptosyringida
Cryptosyringida includes Ophiuroidea, Echinoidea, Holothuroidea, all with closed ambulacra in which the radial nerve is internalized and protected by ossicles (Fig 28-20).
Echinozoa
Echinozoa, sister taxon to Ophiuroidea, consists of the urchins and sea cucumbers. In these echinoderms the oral surface and ambulacra have expanded aborally until they enclose almost the entire body except for a small aboral periproctal region with the anus. A bony ring of ossicles surrounds the pharynx. The hemal system is better developed than in other echinoderms. The tube feet have ossicles.
Echinoidea C
Echinoidea includes about 950 extant species of sea urchins, sand dollars, sea biscuits, heart urchins, and their relatives. The dermal ossicles are thin plates fused to form a rigid, more or less spherical, endoskeletal test. Except for a thin outer epidermis, all the soft anatomy is inside the test. The larva is a bilaterally symmetrical echinopluteus. The test is covered by an abundance of movable spines. Tube feet are the major respiratory organs and the madreporite is aboral.
Urchins may be regular or irregular. Regular urchins are the sea urchins, with radial symmetry, globose nearly spherical bodies, and long spines. Most are epifaunal. Irregular urchinsare sand dollars, sea biscuits, and keyhole, heart, and cake urchins. These are usually infaunal in soft sediments and have a superficial bilateral symmetry superimposed on their radial symmetry. The body is usually flattened and the spines short.
Epifaunal species (regular urchins) possess a feeding apparatus known as Aristotle's lantern, equipped with five strong teeth, used for scraping food from hard substrates. The lantern is reduced in infaunal species (irregular urchins) because most of them are deposit feeders.
Clypeasteroida O
Clypeasteroids are flattened urchins known as sand dollars (Fig 28-42) and sea biscuits (Fig 28-41) adapted for living and moving infaunally in soft sediments. The test is flattened to varying degrees on the oral-aboral axis and a secondary bilateral symmetry is imposed on the underlying radial symmetry. Jaws and a simplified Aristotle’s lantern are present. The respiratory tube feet are arranged in petalloids.
Strongly depressed clypeasteroids are known as "sand dollars" because some of them have a flattened disklike shape that resembles a silver dollar (Fig 28-42). Like other irregular urchins most are infaunal deposit feeders with numerous short spines that are used to burrow through soft sediments. As an adaptation to facilitate movement through sediment, irregular urchins tend to be elongate and slightly streamlined along an axis perpendicular to the oral-aboral axis. This imparts a secondary bilateral symmetry on the typical echinoderm radial symmetry (which is why they are said to be “irregular”). Notice that this is not the same as the bilateral symmetry of holothuroids, in which the anteroposterior axis is the oral-aboral axis.
The aboral ambulacral center of clypeasteroids is at the approximate center of the aboral surface and the oral ambulacral center is at the center of the oral surface. The anus, however, does not coincide with the aboral ambulacral center (it does in regular urchins) but occupies a position on the posterior part of the oral surface or low on the posterior margin of the aboral surface (Fig 28-42). The evolutionary migration of the anus away from its ancestral position at the aboral pole can be followed in fossil echinoids. In contrast, the mouth remains associated with the oral ambulacral center, and for good reason. The small tube feet of the oral ambulacra make up a set of radiating conveyor belts that transport food particles from the periphery to the central mouth (Fig 28-43). No lunules are present.
Laboratory Specimens
Clypeaster rosaceus, the sea biscuit, is a good species for the study of the test of irregular urchins (Fig 28-41A). It is not a good choice for the study of internal anatomy because of the difficulty of opening the thick test. This Caribbean species lives in and near beds of turtle grass on sand in shallow water.
Its body is very thick, unlike those of other sand dollars. The densely matted roots of turtle grass make digging difficult for such a large, bulky animal and it does not burrow. Instead, it uses the short spines of its oral surface to pole its way across the surface of the sand. Clypeaster covers itself with grass blades, shell fragments, and other debris making it very difficult to see in its natural habitat. This camouflage is held in place by suckered tube feet on the aboral surface.
In contrast, its congener, C. subdepressus, does burrow through sediment and is much flatter, being shaped like a conventional, but very large, sand dollar. It lives in areas of open sand free of grass roots.
Clypeaster rosaceus is longer over the aboral-oral axis and narrower on the left-right axis. Its test wall is much thicker. Bilateral symmetry is more evident in C. rosaceus. In comparison,Clypeaster subdepressus is quite thin, presumably for cutting through the sediment. The periproct is on the posterior margin of the oral surface in C. rosaceus but farther in on the oral surface inC. subdepressus.
Cleaned and dried tests of Clypeaster rosaceus and, optionally, C. subdepressus, should be available in the laboratory for study. Devote most of your attention to C. rosaceus but compare it with C. subdepreessus when appropriate.
Test
This exercise is limited to study of the test and does not consider other aspects of sea biscuit anatomy. Examine a dried and bleached test of the sea biscuit, C. rosaceus using the dissecting microscope when needed (Fig 1). Do not use a pen or pencil as a pointer. In life the test is covered by a thin integument and is covered by short, dark brown spines (Fig 28-41A). An unbleached test in which the spines are still intact may be on demonstration. Visit it.
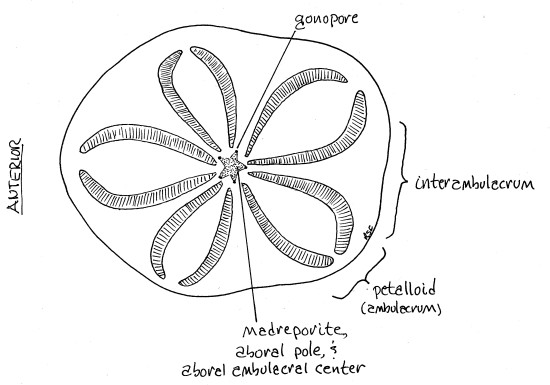
Figure 1. Aboral view of Clypeaster rosaceus. Echinoid50L.gif
Symmetry
The test is a calcareous endoskeleton composed of fused dermal ossicles. It is rigid and, in C. roseaceus, very thick (Fig 1). The convex, upper surface is aboral and the lower, strongly concave, surface is oral. The test is elongated along an axis perpendicular to the oral-aboral axis and this superimposes bilateral symmetry over the original radial symmetry. The bluntly pointed end of the body leads the way when the animal moves and is referred to as anterior. The opposite end, which is flattened, or truncate, is posterior. The plane of bilateral symmetry includes both the antero-posterior axis and the oral-aboral axis. This species is not nearly so flat as most clypeasteroids so the name "sand dollar" is not descriptive. It is usually called a sea biscuit.
Aboral Surface
The aboral surface bears a conspicuous flower-like arrangement of five ambulacral areas called petalloids (= petalloid ambulacra, Fig 1, 2). The term petalloid is an obvious allusion to the petal of a flower. Each petalloid consists of two rows of paired podial pores to accommodate two rows of respiratory tube feet, or podia. Respiratory tube feet have paired pores to accommodate the two connections each tube foot has with its ampulla. Respiratory tube feet are restricted to the aboral surface.
Figure 2. One of the five petalloids of the aboral surface of Clypeaster subdrepressus. Echinoid51L.gif
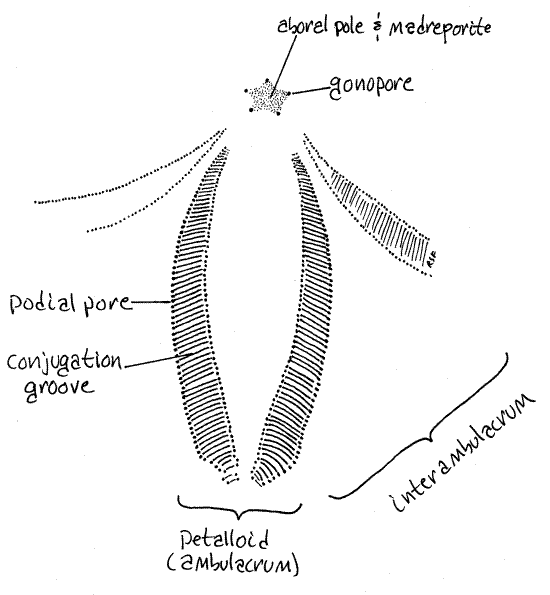
The margins of the petalloid are formed by the paired pores. The two pores of each pair are connected to each other by a shallow conjugation groove. Pore pairs joined by a groove are said to be conjugate.
Examine the margins of a petalloid with magnification and find the two pores of a pair and the groove connecting them. Notice that the groove is shallow and does not penetrate the test but the pores do.
Examine the aboral surface with high power of the dissecting microscope. Most of it is covered by regularly spaced tubercles, each set in a small depression, or pit. In life, the short spines of the animal articulate with these tubercles (Fig 28-31B). The tubercles function as the male half of a ball and socket joint that allows great freedom of movement of the spines (Fig 28-32A). Muscles that operate the spine originate on the test around the circumference of the spine and insert on its base. The test surface between the pitted tubercles is covered with smaller tubercles that do not occupy pits.
In addition to the suckerless respiratory tube feet of the petalloids, conventional suckered tube feet are also present. Clypeaster roseaceus uses them to hold grass blades and shells to its body and to move food particles toward the mouth. With about 35X magnification, search the aboral surface for openings of ordinary, or suckered, tube feet. Hold the surface perpendicular to your line of sight and look closely and with carefully adjusted light in the interiors of the petalloids. Here you may see numerous tiny pores, for the exit of suckered tube feet. They are difficult to see if the light is not shining from the correct angle. The suckered tube feet are most abundant in the interiors of the petalloids or, put another way, in or near the ambulacra, which, of course, is where you would expect tube feet to be.
Aboral Pole
The five petalloids radiate outward from the aboral pole at the center of the aboral surface. This center of radiation of the ambulacra is the aboral ambulacral center. One petalloid points anteriorly, toward 12 on an imaginary clock face, and the other four point to 2, 4, 8, and 10 (Fig 1). The areas between successive petalloid ambulacra are interambulacra. They are not as wide as the petalloids.
An inconspicuous star-shaped madreporite is situated in the aboral ambulacral center at the aboral pole (Fig 1, 2). If you examine its surface with high power of the dissecting microscope, you will see that it is perforated by tiny(!) madreporic pores that, in life, open to the axial canal and stone canal of the water vascular system. It also bears tubercles for the articulation of spines.
At the star-points of the madreporite, one in each interambulacrum, are five conspicuous gonopores through which the gonoducts from the gonads pass
Oral Surface
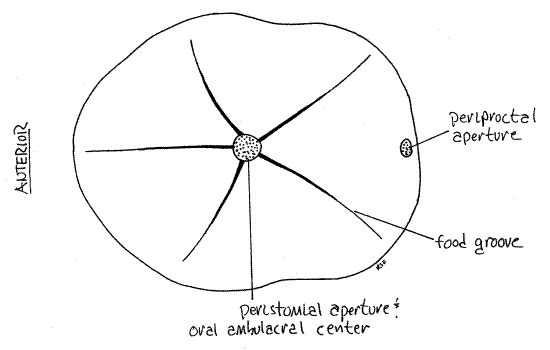
In this species the oral surface is strongly concave with the peristome at the apex of the concavity near the center of the oral surface (Fig 4). The peristome is the region surrounding the mouth (fig 3). In life, the peristome is covered by the peristomial membrane with the mouth at its center, as it is in regular urchins.
The peristome coincides with the oral ambulacral center although the oral ambulacra are not as obvious as the aboral. Five smooth ambulacral food grooves radiate outward from the ambulacral center to the periphery (Fig 3). In position they coincide with the ambulacra, or petalloids, of the aboral surface at 12, 2, 4, 8, and 10 o'clock. Clypeasteroids are deposit feeders. Organic particles are passed by small suckered tube feet along the food grooves to the mouth (Fig 28-43).
The periproct is near the posterior border of the oral surface (Fig 3, 4). In life the anus is located at the center of the periproct which is covered by the periproctal membrane. Note that the periproct and anus are not located at the aboral ambulacral center as they are in regular urchins.
Examine the oral surface with high power of the dissecting microscope. Again you will see pitted tubercles but here they are larger and better developed than they are on the aboral surface. The spines that articulate with these tubercles are the locomotory organs of the biscuit and are stronger than those on the aboral surface. The biscuit uses them to pole its way over the sediment surface.
Use high power of the dissecting microscope to look in the ambulacral areas for podial pores. They are tiny but abundant near the ambulacral food grooves but are scarce in the middle of the interambulacral areas. The oral tube feet have suckers and are used to move food to the mouth.
Figure 3. The oral surface of Clypeaster rosaceus. Echinoid52L.gif
Bisected Test
Examine a test that has been bisected along the median sagittal plane. This is easily accomplished with a hacksaw but the instructor will probably provide you with previously bisected biscuits. Notice the thickness of the wall of the test and find the heavy, oblique pillars that extend between the oral and aboral surfaces to strengthen the test (Fig 4).
Five pairs of erect auricles form a circle, the perignathal ring, around the peristome surrounding the mouth (Fig 4). The auricles insert into deep fossae on the surface of the jaws of Aristotle's lantern and hold the jaws in place.
Study the pieces of Aristotle's lantern if they are present. The lantern, which is the jaw apparatus of urchins, is present among irregular urchins only in clypeasteroids. In these urchins it is large but is much simpler than the lantern of regular urchins (Fig 28-38) and most of the ossicles present in regular urchins are missing. The lantern and teeth of Clypeaster cannot be extended out of the mouth as it is restrained by its articulation with the auricles. The lantern is made of five jaws, each of which supports a hard, glossy, renewable tooth.
Each jaw is composed of two large, winglike, half pyramids (= alveoli). The other ossicles characteristic of the jaws of regular urchins are either absent (e.g. compasses) or vestigial (e.g. rotules and epiphyses). The half pyramids bear two flat surfaces and numerous thin lamellae that resemble the gills of a mushroom. Each pair of half pyramids forms a pyramid (= jaw),which supports one tooth (Fig 28-38). The two half pyramids of a pyramid articulate with each other by the smaller and more irregular of their two flat surfaces.
Figure 4. Median “sagittal” section of Clypeaster rosaceus. Echinoid53L.gif
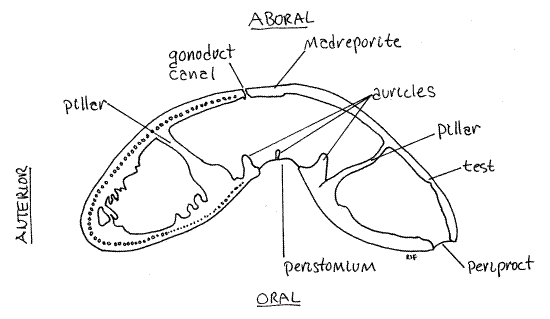
The complete pyramid bears a conspicuous groove on its apex that encloses the tooth. Two deep fossae on the oral surface of the pyramid fit over the auricles of the perignathal ring. The teeth are very hard orally but soft aborally. New tooth material is added aborally as the hard oral end is worn away.
The five pyramids are held together in a pentagonal star surrounding the peristome by comminutor muscles (= interpyramid muscles) which run from the large flat oval surface of one pyramid to the similar opposing surface of the adjacent pyramid (Fig 28-38). Protractor muscles extend from the aboral surface of the pyramids to the auricles of the perignathal ring. In the clypeasteroids these muscles are not able to extend the teeth out of the mouth as they do in the regular urchins. Retractor muscles extend from the inner oral surface of the pyramid to the auricles.
Fit one of the pyramids jaws) in its proper position on an auricle. Then imagine the other four jaws in their positions to get a mental image of the complete lantern.
References
Hyman LH . 1955. The Invertebrates: Echinodermata, vol. IV. McGraw-Hill, New York. 763pp.
Kier PM, Grant RE . 1965. Echinoid distribution and habits, Key Largo Coral reef Preserve, Florida. Smithson. Misc. Coll. 149(6):1-68, 16 pls.
Ruppert EE, Fox RS, Barnes RB. 2004. Invertebrate Zoology, A functional evolutionary approach, 7 th ed. Brooks Cole Thomson, Belmont CA. 963 pp.
Supplies
Dissecting microscope
Teasing needle for use as a pointer
Cleaned test of Clypeaster roseaceus
Cleaned test of C. subdepressus
Bisected test of C. roseaceus