Invertebrate Anatomy OnLine
Zoobotryon verticillatum ©
Ctenostome Bryozoan
5jul2006
Copyright 2001 by
Richard Fox
Lander University
Preface
This is one of many exercises available from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises, a glossary, and chapters on supplies and laboratory techniques are also available at this site. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Lophophorata SP, Bryozoa P, Gymnolaemata C, Eurystomata SO, Ctenostomata O, Stolonifera sO, Vesicularoidea SF, Vesicularidae F (Fig
25-35, 9-26)
Lophophorata SP
Lophophorata includes three taxa, Phoronida, Bryozoa, and Brachiopoda, sharing several morphological characteristics. Some zoologists include Kamptozoa in this group. The first three taxa possess a funnel-shaped anterior ring of ciliated tentacles known as a lophophore (Fig 25-2, 25-25A). The lophophore surrounds the mouth and is an upstream collecting system for suspension feeding. Its tentacles are hollow with extensions of a coelomic space thought to be the mesocoel. The gut is U-shaped with the anterior mouth at the center of the lophophore. The anus is also anterior, but is dorsal to the mouth, outside the ring of the lophophore (Fig 25-2A).
The lophophoral tentacles bear two types of cilia. Frontal cilia on the inside face of the tentacles extend into the interior of the lophophore, whereas lateral cilia on the sides of the tentacles extend into the gap between adjacent tentacles. The mouth is at the bottom (apex) of the lophophore funnel and is encircled by the ring of tentacles. The feeding current is generated by the lateral cilia. In lophophorates and many filter feeding deuterostomes (but not Kamptozoa) water enters the open end of the lophophore, moves toward the mouth, and then exits laterally between the tentacles (Fig 25-25A). Food particles are captured on the upstream side of the tentacles and transported to the mouth by the frontal cilia. In cross section the lophophore may be horseshoe-shaped or circular (Fig 25-4).
Lophophorates inhabit a secreted enclosure, tube, shell, or zooecium that may be organic or mineral. The body is divided into two parts, the mesosome and metasome, each with a coelomic space. The small mesosome is the region of the lophophore and the much larger metasome is the trunk and accounts for most of the body. The tiny anterior epistome is sometimes considered to be a third body region homologous to the protosome of early deuterostomes. Lophophorates are suspension feeders and most are marine but some occur in freshwater.
Bryozoa P
Bryozoa (moss animals) are derived, almost always colonial, lophophorates. Colonies are composed of individuals, or zooids, which are usually less than 0.5 mm in length. Each zooid inhabits a secreted box, the zooecium, into which is can retract. Because of their small size, hemal, excretory, and respiratory systems are absent.
Colonies are usually attached to firm substrata and may be encrusting layers a single zooid thick, or foliose and leaflike, or fruticose (bushlike) and branching (Fig 25-15). Some, such as Zoobotryon, are stoloniferous and consist of tangles of long slender stolons (which are zooids). Bryozoans often resemble seaweeds (or mosses) with which they are frequently confused by the public. Alone among lophophorates, Bryozoa includes freshwater representatives.
The lophophore may be circular or horseshoe shaped in cross section. A small epistome may be present anterior to the mouth. The zooecium can be composed of chitin, calcium carbonate, or gelatinous organic compounds. Many bryozoans are polymorphic with zooids specialized for different functions.
Gymnolaemata C
Gymnolaemate bryozoans are almost entirely marine and have circular lophophores. No epistome and no body wall musculature are present. These are considered to be derived characteristics. Colonies are polymorphic with specialization of zooids.
Ctenostomata O
Ctenostome bryozoans have membranous, gelatinous, or chitinous non-calcified zooecia. The orifice is terminal and has no operculum.
Laboratory specimens
Zoobotryon verticillatum (=Z. pellucidum) is a common, shallow-water, stoloniferous bryozoan. It is a cosmopolitan species known from the southeastern United States, the Mediterranean Sea, Bermuda, California, Hawaii, and elsewhere. It is well suited to laboratory study of Bryozoa because of its transparent body wall through which internal anatomical details are easily seen.
Like other gymnolaemates, Zoobotryon has a circular lophophore and lacks unspecialized body wall muscles. As a ctenostome it has an uncalcified zooecium. Its growth form is stoloniferous but, unlike many stoloniferous species, its branching stolons are erect and extend away from the substratum. The masses of stolons resemble the noodles of oriental cuisine. The colonies may be large, exceeding 15-20 cm in diameter. Zoobotryon is common on ship hulls and may clog power plant or industrial seawater pipes.
The exercise is written for use with fresh, living material but most of the exercise can also be accomplished with preserved or mounted material. The study should be begin with a low power examination using a dissecting microscope followed by higher magnification observation, also with a dissecting microscope.
Anatomy
Use the dissecting microscope on low power to examine a piece of a healthy colony in a 8-cm culture dish of seawater. Observe the overall structure of the colony (Fig 1, 25-15D). It is composed of long, branching, tubular, transparent stolons (kenozooids) which extend above the substratum and to which the feeding individuals, or zooids (autozooids), are attached.
Zooids
The colony is polymorphic (dimorphic really) and is composed of two types of zooids. The vase-shaped autozooids are the feeding zooids (Fig 1, 25-16B). They are typical bryozoan zooids equipped with lophophore, gut, and reproductive equipment.
Figure 1. A small piece of a colony of Zoobotryon verticillatum from the Indian River at Ft. Pierce, Florida. Bryozoa63L.gif
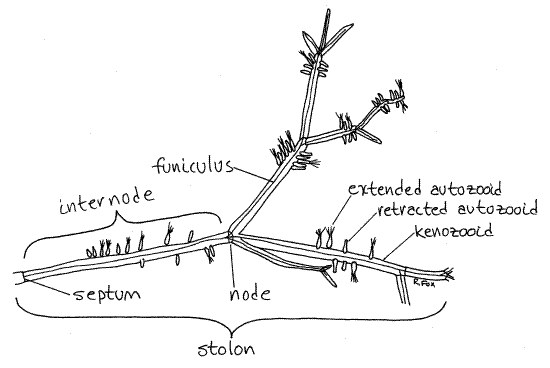
Zooids specialized for some function other than feeding are referred to as heterozooids, of which there may be many types among bryozoans. In ctenostomes, however, the only heterozooids are the kenozooids, of which the stolon is made. Look closely at a stolon and note that it is made of numerous segments, each of which is a kenozooid (Fig 1). Kenozooids have no lophophore or gut and do not feed. They bear little resemblance to autozooids and are much larger. The kenozooids account for most of the biomass of the colony and are easily the most conspicuous part. Note that where kenozooids join each other, the stolon branches. The branching is usually trichotomous but sometimes dichotomous or irregular. The points of branching are nodes (Fig 1). The space between two successive nodes is an internode.
Increase the magnification a little and look at the junction of two kenozooids. Note the partition, or septum, separating the two zooids. The colony may be encrusted with fouling organisms that make it difficult to see the surface or the interior of the zooids. Search the colony, especially the young tips of the stolons, for areas relatively free of fouling.
The epidermis of a bryozoan zooid secretes a nonliving, extracellular cuticle, or zooecium (zoo = animal, oikos = house, thus “animal house”), outside itself. In ctenostome bryozoans, such as Zoobotryon, the zooecium is membranous, flexible, and transparent. In some bryozoans, such as Membranipora and Schizoporella it is calcareous, rigid, and opaque. In others, such as Alcyonidium, it is soft and gelatinous.
Look at a kenozooid and examine its zooecium. The zooecium is underlain by the epidermis, which secretes it, and the mesothelium. Together the zooecium, epidermis, and mesothelium constitute the body wall. There are no unspecialized body wall muscles in gymnolaemate bryozoans. The body wall (zooecium, epidermis, and mesothelium) is known as the cystid to distinguish it from the rest of the zooid, which is called the polypide. The polypide is the part of the zooid not contributing to the body wall and includes the lophophore, gonads, and gut. The spacious interior of the kenozooid is the trunk coelom, or metacoel.
Gently probe the zooecial wall of one of the kenozooids with your minuten nadel (refer to the Techniques chapter for instructions on making these useful tools). Trap the zooid between the nadel and the bottom of the dish so it cannot move. Demonstrate to yourself the flexibility of the zooecium. Note the turgidity of the kenozooid. Use your fine forceps to bend one of the long kenozooids and note again its turgidity. Now push the nadel completely through the body wall into the interior of the coelom. Note the effect of this puncture on the turgidity of the zooid. It looses turgor and becomes flaccid.
Use the forceps again to bend the now collapsed zooid and note the difference. It is more easily bent now than it was when turgid and does not return readily to its original shape. Note the flexibility of the walls under these conditions. What process do you suppose establishes and maintains the turgor in the interior of the kenozooid? The role of the zooecium is similar to that of a plant cell wall in maintaining turgor in plant cells.
Funiculus
Use the highest power available with the dissecting microscope (40-50X) to examine the interior of a kenozooid (Fig 2). Use transmitted light (substage light) to illuminate the interior. Remember that the space within is the coelom but in these specialized zooids few anatomical features are present.
Look carefully in the interior for a slender, transparent thread of tissue extending longitudinally for the length of the zooid. This is the funiculus (Figs 1, 2, 25-23B, 25-16). On close examination it can be seen to be continuous with a network of very long, slender cells extending across the coelom to the body wall and to the basis of the autozooids. The funiculus is composed of similarly long cells.
Look at the septum separating two consecutive kenozooids. In its center is a cluster of yellowish cells known as a rosette (Fig 2, 23E)). The rosette is part of the funicular system and it plugs a pore in the center of the septum. The funiculus attaches to this rosette. The pore is an opening in the septum, reminiscent of plasmodesmata in a plant cell wall. Note that the funiculus divides to send a branch to the rosette of each kenozooid at the node. Note also that the funiculus of the next successive kenozooid attaches to the other side of the rosette. Although the funicular system is not well understood, there is evidence that it is an interzooid transport system by which nutritive material is moved from the feeding autozooids to the non-feeding heterozooids (kenozooids) of the colony. If this is true it would be the analog of the gastrovascular cavity of hydrozoan colonies. The rosettes apparently mediate transport from the funiculus of one zooid, across the septal wall, to the funiculus of the next zooid.
The base of each autozooid joins the kenozooid via a septum similar to that between kenozooids. A rosette is present on each side of this septum also and a branch of the kenozooid funiculus extends to it.
Lophophore
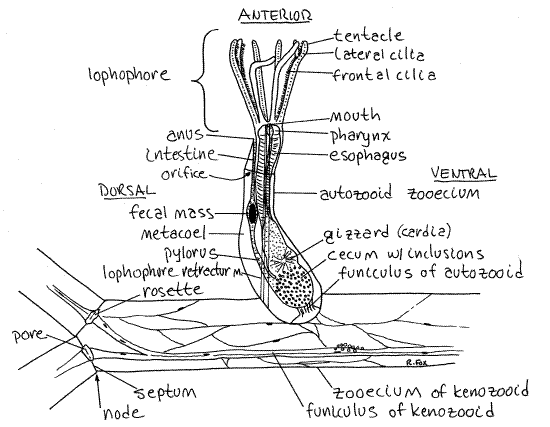
Find some active autozooids with tentacles extended and look at one from the side (Fig 1). The free end of the zooid is anterior and the attached end is posterior. Whereas the kenozooid consists almost entirely of cystid and coelom, most of the autozooid is the polypide, which nearly fills the interior of the zooecium Fig 25-17).
The distal, or free and unattached, end of the zooecium is a circular opening, the orifice, through which the tentacles can be extended. The tentacles are attached to the distal end of the zooid and are borne on a circular ridge of tissue known as the lophophore. (The term lophophore originally referred only to this ridge but is now usually employed in reference to the ridge plus its tentacles.) The region of the lophophore is the mesosome whereas the remainder of the body is the metasome. Like the metasome, the mesosome contains a coelomic space, in this case the mesocoel, or lophophoral coelom. Diverticula of the mesocoel extend into the lophophoral tentacles.
The lophophore and anterior end of the polypide are an introvert that can be extended and retracted. Many of the autozooids in your colony will have retracted lophophores but some should be extended so the tentacles are visible. A pair of lophophore retractor muscles can quickly pull the lophophore and anterior end of the body back into the zooecium. The lophophore must be extended to feed. The most conspicuous features of the autozooid are its zooecium, lophophore, and gut.
Use the nadel to touch gently the tentacles of an extended zooid and watch the response effected by the retractor muscles. Continue watching until the lophophore is reextended. The lophophore is extended by contraction of parietal muscles, which diminish the diameter of the zooid and eject the lophophore (Fig 25-24C, 25-17)
Find an autozooid with the lophophore extended. In Zoobotryon the lophophore has eight tentacles which form an inverted cone with its apex attached to the zooid. The tentacles are hollow with extensions of the mesocoel and are ciliated. The cilia generate the feeding current. The mouth lies at the center of the apex of the cone of tentacles.
>1a. Place a drop of carmine/seawater suspension in the water so there is a cloud of particles in the vicinity of a group of extended zooids. With low power watch the movement of the particles with respect to the lophophore. From which region do particles enter the funnel? Do all particles entering the funnel end up in the gut? Can particles be ejected from the funnel? What are some mechanisms for rejection of particles from the funnel? <
Observe the activity of the cilia on the tentacles. What can you tell of the distribution of these cilia? The inner surfaces of the tentacles bear frontal cilia which are responsible for moving captured food particles toward the mouth. The sides of the tentacles bear lateral cilia, which will be difficult to see if you are looking at the zooid from the side. The lateral cilia generate the feeding current. Describe the direction of the feeding current. Are cilia present on the outside edges of the tentacles?
Figure 2. A single autozooid and part of three kenozooids of Zoobotryon. Bryozoa64La.gif
Bryozoans are noted for the characteristic flicking motion of their tentacles. Observe this motion in one of your animals. It is accomplished by longitudinal muscles in the body wall of the tentacles.
Find an autozooid oriented so you can look straight down into the lophophore and locate the mouth (Fig 2). See if you can see the lateral cilia from this viewpoint.
Digestive System
Now look at an autozooid from the side (Fig 2, 25-17, 25-25A). Most of the tissue you see in the zooid is the gut but it is difficult to distinguish its regions at this magnification.
The gut is U-shaped with both mouth and anus located anteriorly in the vicinity of the lophophore. The mouth opens into a very short muscular pharynx at the base of the lophophore. Watch the pharynx and you should see it dilate occasionally in a process known, appropriately enough, as "gulping" (Fig 25-25C). Dilation is accomplished by contraction of radial muscles running from the pharynx to the body wall. It assists in moving food particles from the lophophore into the gut.
The pharynx opens into a long esophagus, which itself opens into an expanded region known as the stomach (Fig 2). The stomach occupies the bend in the U-shaped gut and consists of three regions; cardia, cecum, and pylorus. The cardia is the descending region, the pylorus ascending, and the cecum in between, in the bend.
In Zoobotryon the cardia is a thick-walled, muscular, spherical gizzard (Fig 2, 25-17). Its epithelium bears tiny denticles, one per cell, which are thought to break the siliceous frustules of diatoms (Fig 25-5D). Zoobotryon can live and grow on a diet of diatoms but bryozoans without a gizzard, such as Bugula, cannot.
Use high power to examine the gut contents looking for diatoms. The cardia opens into the large spacious cecum located at the hairpin bend in the "U" of the gut. From it, the narrow, tubular pylorus exits and ascends to the intestine.
The intestine, with thin walls and narrower diameter, extends anteriorly from the pylorus to the anus (Fig 2, 25-17). The anus is located at the base of the lophophore on the dorsal side of the animal. Ovoid fecal masses may be present in the intestine. The anus is located at the base of the lophophore on the dorsal side of the animal. The anus is outside the ring of the lophophore, as it is in all lophophorates. This position is responsible for the alternative name "Ectoprocta" (=outside anus) for Bryozoa.
Note that the intestine and anus are dorsal, the mouth and lophophore are anterior and the cecum posterior. These are the best landmarks for recognizing the major body axes of these bilaterally symmetric animals. Find anterior, dorsal, posterior, ventral, right and left. Find the plane of symmetry.
Musculature
Gymnolaemate bryozoans lack the unspecialized layers of circular and longitudinal body wall muscles characteristic of phylactolaemates but do have specialized muscles. Well-developed longitudinal muscles are present in the tentacles of all bryozoans. You saw evidence of their presence if you observed tentacular flicking.
Two long lophophore retractor muscles extend from the posterior end of the zooecium to the base of the lophophore (Fig 2, 25-17). These are easily seen in extended autozooids. You watched them operate when you induced the animal to retract earlier in the exercise. Circular parietal muscles are present to pressurize the coelom and extend the lophophore (Fig 25-17).
Retracted Autozooid
Briefly examine a retracted autozooid. Although their orientation with respect to other organs has changed, you should be able to find the retracted lophophore and most of the regions of the gut. When the tentacles are withdrawn, they are enclosed by the retracted body wall (Fig 25-17). The space thus formed contains the tentacles and is the tentacle sheath. In Zoobotryon and some other bryozoans, there is a second chamber distal to the tentacle sheath and similarly enclosed by the retracted body wall. It is the vestibule.
References
Hyman LH. 1959. The Invertebrates. Smaller Coelomate Groups.V . McGraw-Hill, New York. 783 p.
Mukai H, Terakado K, Reed CG. 1997. Bryozoa, pp 45-206 in Harrison FW, Woollacott RM (eds), Microscopic anatomy of Invertebrates, vol 13. Lophophorates, Entoprocta, and Cycliophora. Wiley Liss, New York.
Ruppert EE, Fox RS, Barnes RB. 2004. Invertebrate Zoology, A functional evolutionary approach, 7 th ed. Brooks Cole Thomson, Belmont CA. 963 pp.
Ryland JL. 1970. Bryozoans. Hutchinson Univ. Press, London. 175 p.
Woollacott RM, Zimmer RL. 1977. Biology of Bryozoans. Academic Press, New York. 566 p.
Zirpolo, G. 1933. Zoobotryon verticillatum (Della Chiaje). Mem. Accad. Nuovi Lincei (2)17:109-442.
Supplies
Dissecting microscope
Compound microscope
Living Zoobotryon
Slides, coverslips
Seawater
8-cm culture dish
Carmine-seawater suspension