Invertebrate Anatomy OnLine
Triarthrus eatoni ©
Trilobite
29may2007
Copyright 2001 by
Richard Fox
Preface
This is one of many exercises available from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises can be accessed by clicking on the link to the left. A glossary and chapters on supplies and laboratory techniques are also available through this link. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Arthropoda P, Trilobitomorpha sP, Ptychopariida O
Arthropoda
Arthropoda, by far the largest and most diverse animal taxon, includes chelicerates, insects, myriapods, and crustaceans as well as many extinct taxa such as Trilobitomorpha. The segmented body primitively bears a pair of jointed appendages on each segment. The epidermis secretes a complex cuticular exoskeleton which must be molted to permit increase in size. Extant arthropods exhibit regional specialization in the structure and function of segments and appendages but the ancestor probably had similar appendages on all segments. The body is typically divided into a head and trunk, of which the trunk is often further divided into thorax and abdomen.
The gut consists of foregut, midgut, and hindgut and extends the length of the body from anterior mouth to posterior anus. Foregut and hindgut are epidermal invaginations, being derived from the embryonic stomodeum and proctodeum respectively, and are lined by cuticle, as are all epidermal surfaces of arthropods. The midgut is endodermal and is responsible for most enzyme secretion, hydrolysis, and absorption.
The coelom is reduced to small spaces associated with the gonads and kidney. The functional body cavity is a spacious hemocoel divided by a horizontal diaphragm into a dorsal pericardial sinus and a much larger perivisceral sinus. Sometimes there is a small ventral perineural sinus surrounding the ventral nerve cord.
The hemal system includes a dorsal, contractile, tubular, ostiate heart that pumps blood to the hemocoel. Excretory organs vary with taxon and include Malpighian tubules, saccate nephridia, and nephrocytes. Respiratory organs also vary with taxon and include many types of gills, book lungs, and tracheae.
The nervous system consists of a dorsal, anterior brain of two or three pairs of ganglia, circumenteric connectives, and a paired ventral nerve cord with segmental ganglia and segmental peripheral nerves. Various degrees of condensation and cephalization are found in different taxa.
Development is derived with centrolecithal eggs and superficial cleavage. There is frequently a larva although development is direct in many. Juveniles pass through a series of instars separated by molts until reaching the adult size and reproductive condition. At this time molting and growth may cease or continue, depending on taxon.
Trilobitomorpha
The extinct trilobites are among the earliest known arthropods, appearing first as Cambrian fossils and departing forever in the Permian. Far from being primitive, however, they are already highly derived, specialized arthropods when they make their first appearance and undergo relatively little change over their 300 million year history. These are the best known, to science and the public alike, of fossil invertebrates and are common worldwide.
Although there are eight higher taxa (Fig. 17-7*) and over 15,000 species in 150 families, trilobites exhibit a relatively uniform body plan that includes a heavily calcified dorsal exoskeleton, segmented body with paired, biramous, segmental appendages, and tagmosis consisting of anterior cephalon, middle thorax, and posterior pygidium. The ventral surface is uncalcified and its details, including appendages, rarely fossilize. The body is dorsoventrally flattened, or depressed.
The eponymous three lobes lie side by side and consist of a median axial lobe flanked by a pleural lobe on either side (Fig 17-1). With the exception of the antennae, the apendages are biramous and similar, except for size, over the length of the body. Most are relatively small, 3-10 cm although the full range is 0.5 mm -70 cm.
The appendages have heavy gnathobases whose teeth face, and form, a median food groove. Presumably, movements of the appendages move food particles anteriorly in the food groove to the posterior-facing mouth where ingestion occurs. This primitive arthropod feeding mechanism appears in many other taxa including horseshoe crabs.
The lenses of trilobite compound eyes are mineral, being composed of calcite, rather than being organic as are those of all other arthropods.
A recent interpretation of arthropod phylogeny abandons the name Trilobitomorpha and places trilobites in the taxon Trilobita C. In this scheme Arthropoda P is divided into the sister taxa Schizoramia sP (with biramous appendages and including Crustacea, Trilobita, Chelicerata) and Atelocerata sP (with uniramous appendages and including Hexapoda and Myriapoda). In this revision Trilobita and Chelicerata are sister taxa in Arachnomorpha SC, which is itself the sister taxon of Crustaceomorpha SC within Schizoramia.
Laboratory Specimens
This exercise is based on dorsal and ventral resin casts of Triarthrus eatoni. These casts were made from brass molds prepared from reconstructions by Charles Beecher of the Yale Peabody Museum in 1897. The original reconstructions were based on specimens collected from Upper Ordovician deposits near Rome, NY. The casts are about 2.5 times the actual size of the fossils. Until recerntly they were marketed by Carolina Biological Supply Co. at reasonable cost but unfortunately are no longer available. An encyclopedic treatment of trilobite morphology and identification is provided by Gon (2001).
External Anatomy
Dorsal Aspect
Study the cast of the dorsal surface of Triarthrus using a dissecting microscope as needed. The trilobite dorsum is covered by the calcitic, readily fossilized exoskeleton known as the dorsal shield (Fig 1, 17-1A). The delicate, uncalcified, and rarely fossilized ventral membrane extends across the ventral surface.
The dorsal shield is conspicuously divided into three side-by-side longitudinal lobes. The central, or median, lobe is the axial lobe and is the thickest of the three. In the living trilobite it contained most of the viscera, including, presumably, the gut, heart, and segmentally ganglionated ventral nerve cord (Fig. 17-2A). On either side of the axial lobe lies a thinner pleural lobe. Each pleural lobe is separated from the axial lobe by a longitudinal axial furrow. These three lobes are the basis for the name “ trilobite”.
Tagmata
In addition to the three longitudinal lobes, the body is divided transversely into three sequentialtagma consisting of the anterior cephalon, middle thorax, and posterior pygidium (Fig 1, 17-1A).
Cephalon
The cephalon, or head, bears the sense organs and mouth and is usually relatively large. Internally it housed the brain and anterior gut, including the stomach and digestive ceca (Fig 17-2B). The trilobed construction of the body is readily apparent on the cephalon. The axial lobe extends anteriorly onto the cephalon where it is known as the glabella and the two pleural lobes are present lateral to the glabella. These lateral areas of the cephalon are the genae, or cheeks. A thickened border, the doubleure, surrounds the anterior and lateral margins of the cephalon. A pair of segmented filiform antennae emerge from under the anterior border (Fig 17-1A).
The cephalon is composed of several fused segments but dorsally its segmental nature is not obvious, although some indications of it are evident (Fig. 1, 17-1). The glabella of Triarthrus bears transverse or oblique glabellar furrows that mark the divisions between the four segments of the head.
Figure 1. Dorsal aspect of the Beecher cast of Triarthrus. Appendages have been omitted from the right side.
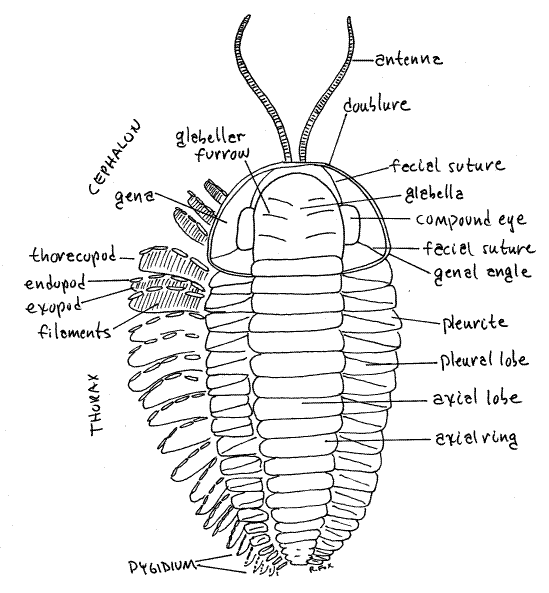
The genae bear a pair of compound eyes. Each eye is composed of numerous ommatidia whose lenses, or facets, are not visible in these casts. In most trilobites the lenses are numerous (up to 15,000 in each eye) and contiguous so that each is hexagonal in shape. A common cornea covers all the lenses of the eye. Such eyes are holochroal. The schizocroal eyes characteristic of some trilobites have larger ommatidia whose circular lenses are widely spaced and do not touch (Fig. 17-2B). Each schizocroal lens is covered by its own cornea. Unlike all other arthropods, trilobites lenses are mineral, made of calcite, rather than organic cuticle> Each lens is a single calcite crystal.
The postero-lateral corner of the gena is the genal angle. In many taxa each of the genal angles bears a genal spine, which may be quite large (Fig. 17-7E,G) but these are absent in Triarthrus.
Of importance in trilobite taxonomy and biology are the facial sutures on the genae. These grooves arise at the anterior margin of the gena and pass posteriorly through the genae, separating the eye from the glabella, and then ending posteriorly. The location of the posterior end of the furrow is taxonomically important. In Fig 1 the suture terminates on the posterior margin of the gena and is opisthoparian. A suture that ends on the lateral margin is proparian and one that ends at the genal angle is gonatoparian. Triarthrus has gonatoparian sutures (Fig 17-1A).
The facial sutures are ecdysial lines along which the old exoskeleton splits during ecdysis. The facial suture divides its gena into a medial fixigena and a lateral librigena (Fig 1). During ecdysis the fixigena remains fused with the glabella, whereas the librigena is liberated from it. The combined fixigena and glabella are known as the cranidium. The molting animal emerges from the exuvium through the opening created by the sutures.
Thorax
The remainder of the body is the trunk, divided into thorax and pygidium, both of which are conspicuously segmented (Fig 1, 17-1A). The thorax, which is the middle region of the body, is much larger than the pygidium. Among trilobites the number of thoracic segments varies from 2-60. How many segments are present in the thorax of your specimen? ______ Thoracic segments are articulated, fore and aft with each other by flexible articular membranes.
Each segment has a central axial ring from which extend, one on the right and one on the left, two pleurites. The combined axial rings of all the segments make up the axial lobe, just as the combined pleurites on each side make up the two pleural lobes. In some trilobites, but not Triarthrus, the pleurites bear lateral spines (Fig 17-7A).
The flexibly articulated segments of the thorax allow for enrollment. In this maneuver the animal arches the body dorsally and brings the ventral surfaces of the pygidium and cephalon in contact with each other (Fig 17-5). This protects the vulnerable ventral membrane and appendages from attack. Most trilobites were able to enroll but some were better adapted for it than others. Triarthrus was capable of complete enrollment and enrolled fossils are common. Some modern arthropods, such as pillbugs and some millipedes, also enroll.
Pygidium
The pygidium is the posteriormost tagma and in Triarthrus it is small and composed of only 4-5 segments visible dorsally (Fig 17-1A). The relative size of the pygidium and cephalon varies with taxon and is an important character in trilobite classification and identification. Like the thorax, the pygidium is composed of appendage-bearing segments but in this case the segments are fused into a rigid platform and are not connected by flexible articular membranes (Fig 1). Although fused, the pygidial segments are distinct. How many pygidial segments are present in your specimen? _____ Pygidial segments and their appendages diminish rapidly in size posteriorly.
The axial lobe extends posteriorly onto the pygidium but tapers to a blunt point posteriorly. The right and left pleural lobes also extend onto the pgidium where they join each other posterior to the end of the axial lobe.
Ventral Aspect
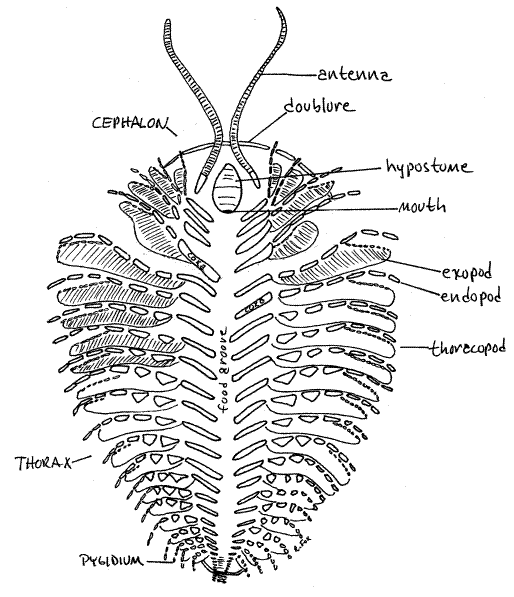
Examine the cast of the ventral aspect of Triarthrus using the dissecting microscope as needed (Fig 2, 17-1B). With the exception of some specific areas, most notably the doublure and hypostome, the ventral surface consists of the delicate, weakly calcified ventral membrane which rarely fossilizes. The segmental appendages are ventral also but, like the ventral membrane, are rarely present in fossils. Happily, one of the unusual features of these Triarthrus fossils is that their appendages are beautifully fossilized.
Figure 2 Ventral surface of the Beecher cast of Triarthrus.
Cephalon
Look at the ventral surface of the cephalon (Fig 2, 17-1B). Along its margins the dorsal shield turns ventrally to form a thick, marginal, ventral shelf of calcified exoskeleton known as the doublure (Fig 2, 17-1B). The doublure typically fossilizes along with the dorsal shield. The doublure of your Triarthrus specimen is a narrow ridge visible only along the anterior margin of the cephalon. Along other borders it is mostly hidden by the appendages.
On the ventral surface, corresponding in position to the glabella, is a longitudinal ridge covered by a sclerite, the hypostome (Fig 2, 17-1B). The hypostome may homologous to the crustacean labrum. Unlike the ventral membrane, the hypostome is calcified and is often present in fossils.
The mouth, which is not visible on your specimen, opens at the posterior edge of the hypostome and faces posteriorly toward the anterior end of the food groove. The gut is J-shaped with the esophagus and stomach located in the central lobe (glabella) of the cephalon (Fig 1, 17-2B). Large digestive ceca extend from the stomach into the genae. The stomach extends farther anterior than any other part of the gut. The esophagus extends posteriorly from the stomach to connect with the backward-facing mouth (Fig 2 17-5B). The gut is a long tube housed in the axial lobe and ends at a posterior ventral anus on the pygidium (Fig 17-2A, 17-3, 17-5B).
The trilobite cephalon is segmented and is probably composed of the acron and four fused segments. Supporting this hypothesis are its four pairs of appendages, which are a pair of antennae and three similar pairs of biramous legs. Each pair of appendages is associated with one of the four cephalic segments.
The antennae are long, filiform, and uniramous. They arise lateral to the hypostome and are the anteriormost appendages (Fig 2, 17-1B). Only one pair is present. The three pairs of postantennal cephalic appendages are biramous and, although they resemble the trunk appendages, they are smaller.
Thorax
In Triarthrus the thorax is by far the largest tagma. Ventrally it is covered by the ventral membrane with which the segmental thoracic appendages, or thoracopods articulate. Most of what you see on the ventral thorax of Triarthrus is appendages. These appendages are biramous and are similar to each other and to the postantennal cephalic appendages.
Each thoracic appendage arises from the ventral membrane near the midline. Its first article is the elongate proximal coxa which can be seen clearly on your specimen. Arising from the lateral end of the coxa are two rami, or branches. The endopod is a leglike stenopod consisting of six cylindrical articles and a terminal claw (Fig 2, 17-1B, 1-2A). It is probably a walking leg. The exopod is a phyllopod consisting of lamella of closely spaced filaments. Its function is not known but it may be a gill for gas exchange, a paddle for swimming or stirring up food particles, or a filter for suspension feeding.
The medial end of each coxa bears a toothed gnathobase, but it is not visible in these specimens (Fig 17-2B). The gnathobases of the right and left coxae of each pair of appendages face each other across the ventral midline. The open space between the coxae is the food groove, which is visible in these casts (Fig 17-1B). Walking or feeding movements of the appendages move food particles into the food groove. The same movements cause the teeth of the gnathobase to grind the food and push it anteriorly in the food groove. Eventually the food arrives at the posteriorly oriented mouth and is ingested.
Pygidium
Ventrally the pygidium is not clearly defined in these preparations.
Internal Anatomy
Little is known of the internal anatomy of these extinct arthropods but some of the information discussed above is available from X-ray studies and the remainder is conjecture based on our knowledge of living arthropods.
Optional Exercise
> a. If fossils or casts of other trilobites are available in the laboratory, study them using your newly acquired knowledge of trilobite morphology. Find their major anatomical features and compare them with Triarthrus. <
*Hyphenated call-outs, such as this one, refer to figures in Ruppert, Fox, and Barnes (2004). Call-outs without hyphenation refer to figures embedded in this exercise.
References
Cisne JL. 1975. Anatomy of Triarthrus and the relationships of the Trilobita. Fossils and Strata 4:45-63.
Fortey R. 2000. Trilobite, Eyewitness to evolution. Vintage Books, New York. 284 pp.
Fortey RA. 1997. Classification, in Kaesler RL (ed.) Treatise on Invertebrate Paleontology, Part O Arthropoda 1, Trilobita, revised. Vol. 1: Introduction, Order Agnostida, Order Redlichiida. Geological Society America & Univ. Kansas, Boulder, CO. 530pp.
Fortey R. 2000. Trilobite: Eyewitness to evolution. Vintage Books, New York. 284 pp.
Fortey RA. 2004. The lifestyles of the trilobites. Am. Sci. 92(5):446-453.
Gon S. 2001. A pictorial guide to the orders of trilobites. Privately published and available for $15 from S. Gon, 1604 Olalahina Pl, Honolulu, HI, 96807. 90 pp.
Gon S. A guide to the orders of trilobites. www.aloha.net/~smgon/ordersoftrilobites.htm
Ruppert EE, Fox RS, Barnes RB. 2004. Invertebrate Zoology, A functional evolutionary approach, 7 th ed. Brooks Cole Thomson, Belmont CA. 963 pp.
Supplies
resin cast of Triarthrus dorsal aspect*
resin cast of Triarthrus ventral aspect*
*Resin casts of Triarthrus are no longer available from Carolina Biological Supply Company (ER GEO5800)