Invertebrate Anatomy OnLine
Thelyphonus ©
Whip Scorpion
19jun2006
Copyright 2004 by
Richard Fox
Lander University
Preface
This is an exercise from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises can be accessed by clicking on the links to the left. A glossary and chapters on supplies and laboratory techniques are also available. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Arthropoda P, Chelicerata sP, Euchelicerata, Arachnida C, Lipoctena, Megoperculata, Uropygi O, Thelyphonida, Thelyphonidae F (Fig 16-15, 18-44, 18-47)
Arthropoda P
Arthropoda, by far the largest and most diverse animal taxon, includes chelicerates, insects, myriapods, and crustaceans as well as many extinct taxa such as Trilobitomorpha. The segmented body primitively bears a pair of jointed appendages on each segment. The epidermis secretes a complex cuticular exoskeleton which must be molted to permit increase in size. Extant arthropods exhibit regional specialization in the structure and function of segments and appendages but the ancestor probably had similar appendages on all segments. The body is typically divided into a head and trunk, of which the trunk is often further divided into thorax and abdomen.
The gut consists of foregut, midgut, and hindgut and extends the length of the body from anterior mouth to posterior anus. Foregut and hindgut are epidermal invaginations, being derived from the embryonic stomodeum and proctodeum respectively, and are lined by cuticle, as are all epidermal surfaces of arthropods. The midgut is endodermal and is responsible for most enzyme secretion, hydrolysis, and absorption.
The coelom is reduced to small spaces associated with the gonads and kidney. The functional body cavity is a spacious hemocoel divided by a horizontal diaphragm into a dorsal pericardial sinus and a much larger perivisceral sinus. Sometimes there is a small ventral perineural sinus surrounding the ventral nerve cord.
The hemal system includes a dorsal, contractile, tubular, ostiate heart that pumps blood to the hemocoel. Excretory organs vary with taxon and include Malpighian tubules, saccate nephridia, and nephrocytes. Respiratory organs also vary with taxon and include many types of gills, book lungs, and tracheae.
The nervous system consists of a dorsal, anterior brain of two or three pairs of ganglia, circumenteric connectives, and a paired ventral nerve cord with segmental ganglia and segmental peripheral nerves. Various degrees of condensation and cephalization are found in different taxa.
Development is derived with centrolecithal eggs and superficial cleavage. There is frequently a larva although development is direct in many. Juveniles pass through a series of instars separated by molts until reaching the adult size and reproductive condition. At this time molting and growth may cease or continue, depending on taxon.
Chelicerata sP
Chelicerata is a large taxon that includes spiders, scorpions, pseudoscorpions, ticks, mites, horseshoe crabs, sea spiders, and many others. The group originated in marine habitats but almost all modern chelicerates are terrestrial.
The body is divided into an anterior cephalothorax with six pairs of appendages and a posterior abdomen which, in most groups, does not bear appendages or has highly modified appendages. The first appendages of the cephalothorax are the chelicerae. Antennae are not present and the brain has two regions rather than the three found in mandibulates. Appendages are primitively biramous but are uniramous in almost all Recent taxa.
Euchelicerata
The segments of the cephalothorax are fused and covered by a dorsal shield, or carapace. Two median eyes are present.
Arachnida C
Arachnids are the terrestrial descendents of the early aquatic chelicerates. The taxon includes the mites, scorpions, pseudoscorpions, spiders, harvestmen, and several other taxa. All are adapted for a terrestrial existence with internal gas exchange and a waterproof integument. Some are capable of silk production and many use toxins to subdue or kill the prey. Most are carnivores, digestion is usually outside the body, and food is liquefied before ingestion.
Megoperculata
Megoperculata, which consists of Uropygi, Amblypygi, and Araneae, is characterized by biarticulate chelicerae, which contrasts with the ancestral triarticulate condition characteristic of scorpions and apulmonate arachnids (palpigrades, pseudoscorpions, harvestmen, mites, tick spiders, and solifugids). The megoperculate sperm axoneme has a 9 x 2 + 3 microtubule arrangement. In modern phylogenies Schizomida is included in Uropygi but in the past it has often been considered to be separate. When Schizomida sO is placed in Uropygi O, the whip scorpions are assigned to the sister taxon Thelyphonida sO. In some older classifications Uropygi, Schizomida, and Amblypygi, all of which have greatly enlarged pedipalps, are grouped together in a taxon known as Pedipalpi.
Uropygi O
The small, tropical and subtropical taxon Uropygi O comprises two higher taxa, Thelyphonida sO and Schizomida sO. Thelyphonida, the whip scorpions, includes 106 known extant species in 16 genera in one family (Thelyphonidae F). Thelyphonus, with 25 species, is the largest and most widespread genus. Schizomida contains about 180 species of small interstitial chelicerates. Schizomida is sometimes considered to be a separate order. This exercise is devoted to thelyphonidans and does not consider schizomidans.
Figure 1. Dorsal view of the whip scorpion, Thelyphonus. Abdominal tergites are numbered. Unsclerotized integument is stippled. Uropygid16L.gif
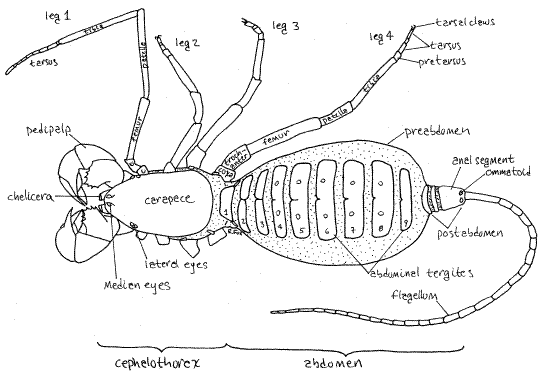
Common names for thelyphonidans include whip scorpion, vinegaroon, and vinegrillo. Whip scorpions are large and robust, in contrast with the small, delicate schizomidans. The pedipalps are massive and may be prehensile. A pair of anal glands in the posterior preabdomen produce a repugnatorial secretion of acetic and caprylic acids which is sprayed, with considerable accuracy, at would-be predators (see Eisner, 2002). The abdomen is divided into a large preabdomen and much smaller postabdomen, both of which are conspicuously segmented. The postabdomen bears a long, slender flagellum (= telson). No poison glands or silk glands are present.
Mastigoproctus is an American genus found from southern North America (including the southern United States) to northern South America. It is the only genus found in the United States. Thelyphonellus and Amauromastigon are South American. Etiennius is the only African genus. Thelyphonus is one of 13 genera in southeast Asia. Whip scorpions are absent from Australia and Europe. African uropygidans are thought to be introductions. Whip scorpions are nocturnal predators that prey on other arthropods. Most are cryptic, hiding under rock or logs in moist habitats during the day. A few, including Mastigoproctus, occur in arid regions. Mastigoproctus giganteus, the largest whip scorpion, reaches 8 cm in length (not counting the flagellum).
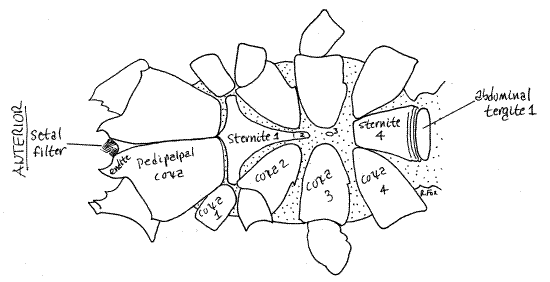
Figure 2. Ventral view of the whip scorpion, Thelyphonus. Abdominal sternites are numbered. Unsclerotized integument is stippled. Uropygid17L.gif
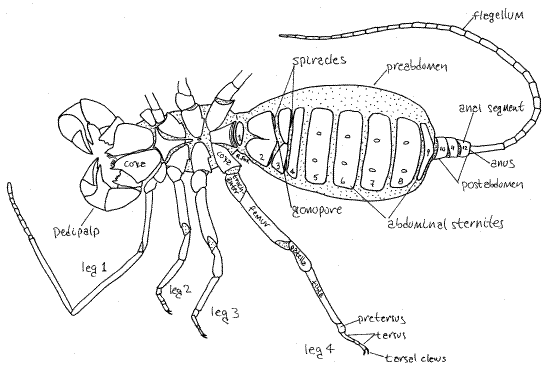
The body consists of a cephalothorax and abdomen separated by a slightly narrower pedicel. The cephalothorax is covered dorsally by a single sclerotized carapace which bears a pair of median eyes and 3-5 pairs of lateral eyes. Ventrally the cephalothorax has three sternites. Chelicerae are small and biarticulate, the distal article being a fang that folds against a tooth on the basal article. The raptorial pedipalps are unusually large. The first legs are long and tactile and are used as sensory antennae. The remaining three pairs of legs are ambulatory and are more robust than the antenniform first legs. The 12-segmented abdomen is divided into the heavy, thick 9-segmented preabdomen and the small 3-segmented postabdomen. A slender whip-like flagellum (often referred to as the “telson”) extends posteriorly from the postabdomen.
The segmented heart may be confined to the abdomen or it may extend into the cephalothorax. Gas exchange is accomplished by two pairs of book lungs with spiracles on the second and third abdominal segments. Coxal glands (two pairs), nephrocytes, and Malpighian tubules are responsible for excretion. The coxal glands open through pores on the coxal of the first and third pairs of legs. The ganglia of abdominal segments 8-12 are fused in the eighth abdominal segment. Vinegaroons are gonochoric with slight sexual dimorphism. Following a courtship ritual the male uses a modified pedipalpal finger to push a spermatophore into the female gonopore on abdominal segment 2. The female releases fertilized eggs into a membranous sac attached to her gonopore. The eggs and embryos are brooded in this sac and upon hatching are carried on the female’s back for a few weeks. Whip scorpions have four juvenile instars, each lasting about a year. Molting ceases after the fourth molt produces the adult.
Laboratory Specimens
Mastigoproctus giganteus is the only whip scorpion found in the United States where it occurs in Florida and the southwest. It is not available commercially. The southeast AsianThelyphonus is available preserved from Ward’s Natural Science. This exercise is limited to external anatomy.
External Anatomy
Tagmata
Typical of arachnids, the thelyphonidan body comprises two tagmata. The anterior tagma is the cephalothorax (= prosoma) which bears the usual six pairs of chelicerate appendages (Fig 1,2,18-16A). Its segmentation is inconspicuous. It is longer than wide and dorsoventrally depressed. The posterior tagma is the abdomen (= opisthosoma) which is conspicuously segmented but has no appendages.
Figure 3. Lateral view of the left side cephalothorax of the whip scorpion, Thelyphonus. This view shows the lateral surface of the unextended left pedipalp. Compare with Fig 6. uropygid18L.gif
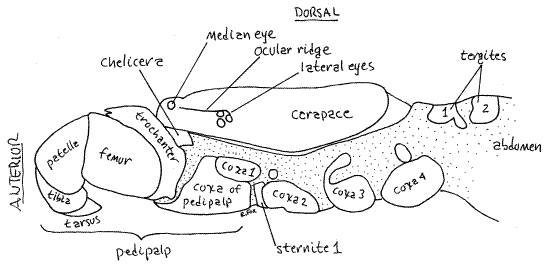
Cephalothorax
The cephalothorax is covered dorsally by a one-piece sclerotized carapace which shows no signs of segmentation (Fig 3,4,18-16A). Ventrally, however, segmentation is apparent in the placement of appendages and sternites. Each of the six segments of the cephalothorax bears a pair of appendages, which are, in order from anterior to posterior, chelicerae, pedipalps, and four pairs of legs
Eyes
Dorsally the carapace of Thelyphonus bears a single pair of median eyes on a low tubercle at the anterior tip of the carapace (Fig 3,4). A cluster of three lateral eyes is located on the margin of the carapace at about the level of the first legs. The number of lateral eyes varies with taxon. In some taxa a low lateral ocular keel, or ridge, extends along the margin of the carapace between the median and lateral eyes of each side.
Figure 4. Dorsal view of the cephalothorax of the whip scorpion, Thelyphonus. Uropygid19L.gif
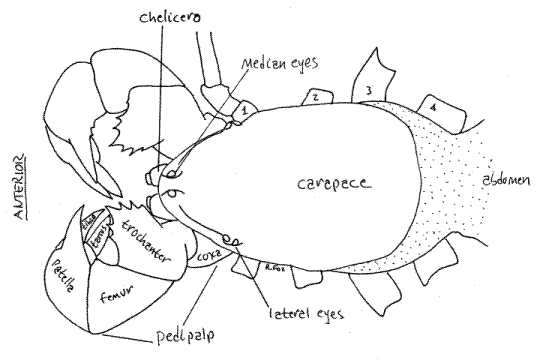
Sternum
Ventrally the cephalothorax is mostly covered by the sclerotized sternites of its segments and the coxae of its appendages (Fig 2,5). The largest, most conspicuous structures of the ventral cephalothorax are the coxae of the two pedipalps. The remainder of the ventral cephalothorax is covered by sternites surrounded by soft, unsclerotized, articulating membranes. The sternites of the four leg-bearing segments are known collectively as the sternum. Sternite 1 is a large, narrowly shield-shaped sclerite between the coxae of the first two pairs of legs. Sternite 2 is a small process on the posterior end of sternite 1. Sternite 3 is very small and is scarcely discernable on the midline between the coxa of the third legs. Sternite 4 is a relatively large trapezoidal sclerite between the two fourth legs.
Appendages
Each cephalothoracic segment bears a pair of appendages typical of chelicerates. These consist of chelicerae, pedipalps, and four pairs of legs, of which the first pair are modified for sensory reception.
Chelicerae
In Thelyphonus the chelicerae are small and are largely hidden by the massive pedipalps, making them difficult to visualize without further dissection (Fig 4). Such dissection, if it is undertaken, is destructive and should be postponed until all other aspects of the external anatomy have been completed.
Figure 5. Ventral view of the cephalothorax of Thelyphonus. The sternites are numbered. Uropygid20L.gif
Thelyphonidan chelicerae are biarticulate, like those of spiders (Fig 6). Many of the features described below will not be apparent without further dissection, as described at the end of this exercise. The two articles are a large, basal, proximal article and a distal movable article that closes against the proximal article to form a pincer. The chelicerae are heavily setose distally. The proximal article has a sclerotized immovable finger on its antero-ventral corner. This finger has two teeth (bifid). The distal article is a movable finger, or fang, that closes against the immovable finger to form a grasping mechanism. In dorsal view, the large proximal article is seen protruding anteriorly from under the anterior margin of the carapace. The movable finger (= fang) extends ventrally from the proximal article and moves in the vertical plane. Hold the specimen vertically so you can focus on the anterior end of the cephalothorax (en face view) while you hold the pedipalps aside. Find the chelicerae and the two opposing fingers of each chelicera. Both fingers of each chelicera are dark, heavily sclerotized and appear fang-like so, in en face view, it appears that this arachnid has four fangs. Only the movable finger, however, is a fang so there are only two. Unlike spiders, whip scorpions have no poison glands.
Pedipalps
The pedipalps dominate the uropygidan cephalothorax (Fig 1,2,3,4,5,6,18-16A). Each consists of six articles which are, in order from proximal to distal, the coxa, trochanter, femur, patella, tibia, and tarsus. The pretarsus (= metatarsus, basitarsus), present in the walking legs, is fused with the tarsus.
The large pedipalpal coxae abut each other and are fused on the ventral midline (Fig 5). Each has an anterior median endite which bears a distal tooth and a dorsal brush of fine setae. The relatively short trochanter bears a dorsal semicircular row of numerous strong teeth which, when in opposition to those of the trochanter of the opposite side, are used to masticate food (Fig 4). The femur is relatively simple, with few teeth. The patella has a large patellar finger extending anteriorly from its medial border. The tibia and tarsus together form a prehensile pincer. The tibia bears a large tibial finger, similar to that of the patella. The tarsus in its entirety is a movable tarsal finger that closes against the tibial finger to form a pincer. The patellar finger also opposes the tibia and tarsus and forms a second pincer with the combined tibia and tarsus serving as the movable finger. The pedipalps are sometimes sexually dimorphic with the patellar finger being longer in males than in females. The endites are fused along the midline to form a trough which, as the floor of the preoral cavity, contains liquefied food on its way to the mouth.
Figure 6. Medial view of the right chelicera and pedipalp of Thelyphonus viewed from the left. The left chelicera and pedipalp have been removed to reveal the preoral cavity. The pedipalp is hyperextended. Compare with Fig 3. uropygid21L.GIF
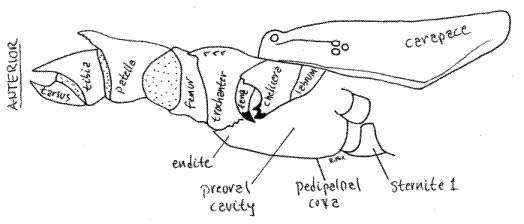
Preoral Cavity
A preoral cavity formed chiefly by the pedipalpal coxal endites precedes the mouth (Fig 6). The cavity will not be seen until later, after removal of the left pedipalp and chelicera. The chelicerae and labrum enclose the cavity dorsally and the fused trough-like pedipalpal coxae form its floor and posterior wall. The arachnid labrum (= upper lip) is an anterior, median, lightly sclerotized region of the body wall overhanging the mouth posterior to the chelicerae and between the pedipalps. No labium is present, it being functionally replaced by the pedipalpal coxae. A setal filter of dense setae on the pedipalpal coxal endites and chelicerae strain the food and exclude all but the smallest particulates from the gut.
Legs
The remaining appendages are four pairs of so-called walking legs, although only three pairs are used for walking. In contrast with the pedipalps, the unmodified legs comprise seven articles (coxa, trochanter, femur, patella, tibia, pretarsus, and tarsus, plus a tarsal claw).
The first legs are slender tactile organs composed of six articles (Fig 1,2,18-16A). The coxa and trochanter are short cylinders extending laterally from the lateral arms of the sternite of cephalothoracic segment 3. The femur is a much longer cylinder and the patella and tibia are longer still. The tarsus is antenniform, being divided into 8-9secondary articles (= tarsomeres). Pretarsus and tarsal claws are absent. These sensory legs are held in front of the animal, antenna-like, as it moves forward. In females of some genera, including Thelyphonus, the tarsal secondary articles of the first legs are swollen or otherwise modified. The first legs are the longest of the four pairs.
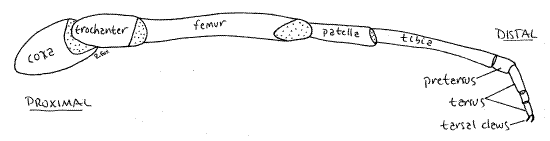
Legs 3-4 are relatively unspecialized, ambulatory legs which lack the specialized antenniform tarsus (Fig 7,18-16A). Each ambulatory leg consists of a large coxa embedded in the venter of the cephalothorax and extending laterally from a sternite. The trochanter is a short cylinder extending between the coxa and the larger, cylindrical femur. The patella, tibia, pretarsus, and tarsus are reduced in diameter. Of them, the tibia is the longest. The tarsus comprises three secondary articles and terminates in a tarsal claw consisting of two hooks. The pretarsus is a short article between the tibia and the proximal tarsal article.
Figure 7. The left leg 4 of Thelyphonus. Articular membranes are stippled. Uropygid22L.gif
Abdomen
The large abdomen consists of 12 segments, of which 1-9 form the large, conspicuous, ovoid preabdomen and 10-12 the small, relatively inconspicuous, cylindrical postabdomen (= pygidium) (Fig 1,2,18-16A).
Preabdomen
Abdominal segment 1 forms the slightly narrowed pedicel, or waist, connecting the abdomen with the cephalothorax . Dorsally the preabdomen bears nine tergites, of which the first is undivided and the remaining eight are divided into right and left portions (Fig 18-16A). The median separation of tergites varies with taxon. Ventrally all nine preabdominal segments have sternites although that of the first segment (pedicel segment) is small and partly hidden. The sternites are short and wide, resembling the tergites, and may or may not be divided into right and left portions, depending on taxon. Sternites and tergites join each other laterally via an extensive pleural membrane.
Abdominal sternite 2, the genital sternite, is large and bears along its posterior border the two spiracles of the first pair of book lungs (Fig 2). The median gonopore is located on the posterior border of this sternite. The spiracles of the second pair of book lungs are on the posterior border of the abdominal sternite 3. In Thelyphonus sternite 3 bears a small posterior median projection, or tooth, absent in females.
Postabdomen
The three segments (abdominal segments 10, 11, 12) of the postabdomen are small and short (Fig 1,2, 18-16A). The postabdominal segments, like those of scorpions, are cylinders formed of fused tergites and sternites with no flexible articulating pleura. The posteriormost segment of the postabdomen (segment 12) is the anal segment, which bears the anus and openings of the two defensive anal glands. The anal segment is longer than the other two postabdominal segments.
The flagellum is also borne on the anal segment (Fig 1,2, 18-16A). The flagellum is often lost or detached in preserved specimens. The long antenniform flagellum is probably homologous to the telson, or sting, of scorpions. It extends posteriorly from the postabdomen and is composed of 30-40 “segments”, which, being post anal, are not homologous to body segments. The anal segment of Thelyphonus has a pair of pale dorsal spots known as ommatoids, reputed, but not demonstrated, to have a photoreceptive function. Some uropygidans have two pairs of ommatoids. The anus is a transverse slit ventral to the base of the flagellum. The anus is best viewed by holding the specimen with its head down while focusing on the distal end of the anal segment. The slit-like anus can be demonstrated with a minuten nadel. The two inconspicuous anal gland apertures are lateral to the anus, one on either side of it. The flagellum may be light sensitive.
Mouthparts Revisited
Once your study of external anatomy is completed you may wish to return to the cephalothorax and make a more detailed study of the chelicerae, pedipalps, and preoral cavity. Doing so requires removal of the left pedipalp and chelicera (Fig 6).
Use a pair of fine (iridectomy) scissors to cut medially between the two pedipalpal coxae and then cut the muscles holding the left pedipalp in place. Remove the left pedipalp. Removal of the this pedipalp will reveal the left chelicera and its structure can be seen to be as described earlier in the chelicerae section. You may study the left chelicera if desired or, if you wish to reveal the preoral cavity, you can use the fine scissors to remove the left chelicera. If you do so, then the preoral cavity will be open to view and the right chelicera and right pedipalp will be intact and can be studied. Use the description in the chelicera section to guide you through this study. The details of the cheliceral pincer mechanism will be difficult to see through the dense setation of the chelicera. This is part of the setal filter that excludes particulates from the arachnid gut. Remove most of these setae with fine forceps and/or iridectomy scissors.
Return to the earlier discussion of the chelicerae, pedipalps, and preoral cavity and find the structures you could not see prior to removal of the mouthparts.
References
Ax P. 2000. Multicellular animals, II. Springer, Berlin. 396pp.
Eisner T. 2002. For love of insects. Belknap Press, Harvard University, Cambridge. 448 pp.
Harvey MS. 2002. The neglected cousins: What do we know about the smaller arachnid orders? J. Arachnology 30:357-372.
Kaestner A. 1968. Invertebrate zoology, Arthropod relatives, Chelicerata, Myriapoda. Wiley Interscience, New York. 472 pp.
Levi HW. 1982. Uropygi, p. 75 in Parker SP (ed.), Synopsis and classification of living organisms, vol. 2. McGraw-Hill, New York. 1232 pp.
Piel W. Arachno Web. www.arachnology.org/Arachnology/Arachnology.html
Rowland JM, Cooke JAL. 1973. Systematics of the arachnid order Uropygida (= Thelyphonida). J. Arachnology 1:55-71.
Ruppert EE, Fox RS, Barnes RB. 2004. Invertebrate Zoology, A functional evolutionary approach, 7 th ed. Brooks Cole Thomson, Belmont CA. 963 pp.
Savory T. 1977. Arachnida, 2 nd ed. Academic Press, New York. 340 pp.
Schultz JW. 1990. Evolutionary morphology and phylogeny of Arachnida. Cladistics 6:1-38.
Vanuytven H. Arachnology Home Page : www.arachnology.be/Arachnology.html
Weygoldt P. 1970. Courtship behaviour and sperm transfer in the giant whip scorpion, Mastigoproctus giganteus (Lucas) (Uropygi, Thelyphonidae). Behaviour 36:1-9.
Weygoldt P. 1971. Notes on the life history and reproductive biology of the giant whip scorpion, Mastigoproctus giganteus (Uropygi, Thelyphonidae) from Florida. J. Zool. Soc. London 164:137-147.
Weygoldt P. 1996. Chelicerata, pp. 449-497 in Westheide W, Reiger RM (Eds.) Spezielle Zoologie. Teil 1. G. Fischer Verlag, Stuttgart. 909 pp.
Supplies
Dissecting microscope
Dissecting set with microdissecting tools
Living or preserved whip scorpion