Invertebrate Anatomy OnLine
Terebratella ©
Articulate Brachiopod
with notes on Terebratalia andTerebratulina
5jul2006
Copyright 2001 by
Richard Fox
Lander University
Preface
This is one of many exercises available from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises, a glossary, and chapters on supplies and laboratory techniques are also available at this site. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Lophophorata SP, Brachiopoda P, Articulata C, Terebratulida O, Terebratellidina sO, Terebratellidae F (Fig 25-35, 9-26)
Lophophorata SP
Lophophorata includes three taxa, Phoronida, Bryozoa, and Brachiopoda, sharing several morphological characteristics. Some zoologists include Kamptozoa in this group. The first three taxa possess a funnel-shaped anterior ring of ciliated tentacles known as a lophophore (Fig 25-2, 25-25A). The lophophore surrounds the mouth and is an upstream collecting system for suspension feeding. Its tentacles are hollow with extensions of a coelomic space thought to be a mesocoel. The gut is U-shaped with the anterior mouth at the center of the lophophore. The anus is also anterior, but is dorsal to the mouth, outside the ring of the lophophore (Fig 25-2A).
The lophophoral tentacles bear two types of cilia. Frontal cilia on the inside face of the tentacles extend into the interior of the lophophore, whereas lateral cilia on the sides of the tentacles extend into the gap between adjacent tentacles. The mouth is at the bottom (apex) of the lophophore funnel and is encircled by the ring of tentacles. The feeding current is generated by the lateral cilia. In lophophorates and many filter feeding deuterostomes (but not Kamptozoa) water enters the open end of the lophophore, moves toward the mouth, and then exits laterally between the tentacles (Fig 25-25A). Food particles are captured on the upstream side of the tentacles and transported to the mouth by the frontal cilia. In cross section the lophophore may be horseshoe-shaped or circular (Fig 25-4).
Lophophorates inhabit a secreted enclosure, tube, shell, or zooecium, that may be organic or mineral. The body is divided into two parts, the mesosome and metasome, each with a coelomic space. The small mesosome is the region of the lophophore and the much larger metasome is the trunk and accounts for most of the body. The tiny anterior epistome is sometimes considered to be a third body region homologous to the protosome of early deuterostomes. Lophophorates are suspension feeders and most are marine but some occur in freshwater.
Brachiopoda P
Brachiopods are filter feeding, marine, benthic lophophorates in two higher taxa, Inarticulata and Articulata. Brachiopoda is ancient and has a rich fossil record of over 12,000 species although fewer than 350 are living today.
The body, consisting of mesosome and metasome, is enclosed in a calcareous or partly organic bivalve shell with dorsal and ventral valves. The body wall is folded to form a mantle composed of dorsal and ventral lobes enclosing a mantle cavity. The body is enclosed in a shell comprising dorsal and ventral valves secreted by the mantle. A fleshy pedicle attaches the animal to the substratum.
The lophophore, which is the feeding and respiratory organ, is large and coiled into two spiral rows of tentacles. The gut is complete in Inarticulata but ends blindly in Articulata. Excretion is via a pair of large metanephridia. The nervous system consists of a circumesophageal nerve ring from which arise nerves to the body. The hemal system includes heart, blood channels, and open sinuses. Brachiopods are gonochoric and fertilization is external.
Articulata C
Articulate brachiopods have a complex hinge mechanism with interlocking teeth and sockets but with a simple musculature. No chitin is present in the periostracum and the shell is calcareous. The dorsal valve is usually equipped with calcareous brachidia to support the lophophore. The gut is blind ending, without an anus. Most live attached to the substratum
Laboratory Specimens
Preserved articulate brachiopods provided by commercial suppliers in North America are usually Terebratella, Terebratulina septentrionalis (Terebratulida O, Terebratulidina sO, Cancellothyridacea SF, Cancellothyridae F), and Terebratalia transversa (Terebratulida O, Terebratellidina sO, Laqueidae F).
Biological supply houses tend to ship these species indiscriminately with inappropriate labels, sometimes with more than one genus in a jar. Terebratella, the species for which this exercise is specifically written, is a Pacific coast animal with a radially ridged shell and a strong septum on the dorsal valve. Terebratulina septentrionalis occurs on the northeastern North American coast and in northern Europe (south to Cape Cod and Norway, respectively). It also has a ridged shell but lacks a septum on the dorsal valve. Terebratalia transversa is a northern Pacific species occurring from Alaska to southern California. It has a smooth shell. Specific, or even generic, identification of your specimen is not really important, but if you do not have Terebratella, you may expect some slight variation between the written descriptions and your specimen.
The dissection can be performed on living or preserved material. Living specimens can be dissected relaxed in isotonic magnesium chloride. All dissection should be conducted under an appropriate liquid in a small dissecting pan on the stage of the dissecting microscope. The liquid should be magnesium chloride for dissection of relaxed specimens or tapwater for preserved specimens.
External Anatomy
Shell
Begin your study of articulate brachiopods with a careful examination of an empty, cleaned shell. If empty shells are not available, postpone the study of the inside of the shell until you have finished with the soft anatomy and can remove it from the shell.
The shell of articulates is calcareous, hard, and rigid. It is composed of two parts, or valves, and is thus bivalve, like that of bivalve molluscs (Fig 3, 25-9). The convergence between the shells of brachiopods and bivalves is remarkable but the differences between them are substantial. Brachiopod valves are dorsal and ventral whereas those of bivalves are right and left. The plane of bilateral symmetry of a brachiopod divides each valve into more or less equal right and left sides whereas the plane of symmetry of a bivalve mollusc divides the shell into right and left valves (which may or may not be equal).
The valves are convex and bulge outward around the animal. The dorsal valve is the smaller, and often the flatter, of the two and it acts as a lid, or cover (Fig 3, 25-9E,G). It is sometimes called the brachial valve because it supports the brachia, or arms, of the lophophore.
The ventral valve is the larger and is usually the deeper of the two. It contains most of the body. It is sometimes called the pedicular valve because it contains a large posterior opening, the aperture (= foramen), through which a tough fleshy stalk, the pedicle, protrudes (Fig 25-8A,C). If you are studying a cleaned shell in which there is no animal, the pedicle will not be present but the aperture will. Find the semicircular foramen and use it to determine which end is posterior. The valves separate from each other, or gape, along their anterior margins. The opening thus formed is the gape (Fig 3, 25-9E).
Use the landmarks above to find anterior, posterior, dorsal, ventral, right, and left. Find the plane of symmetry of these bilaterally symmetrical animals. In the life position the dorsal valve is often down and the ventral valve is up (Fig 25-9C).
The shell, like that of molluscs, is composed of three layers. Outermost is a thin, periostracum composed of protein. It may be visible as a thin, brownish layer on the outside of the valves. It is often worn away, especially on the higher areas of the shell where it is exposed to abrasion. (It will not be present in shells that have been cleaned with bleach.) The remaining two layers are chiefly calcium carbonate. The first of these, and the one in contact with the periostracum, is the primary layer consisting of granular calcium carbonate (Fig 25-8D). The secondary layer is innermost and is composed of calcium carbonate and protein. The shell is secreted by the underlying epidermis, known as the mantle. The edge of the mantle secretes the periostracum and the primary layer whereas the entire outer mantle surface secretes the secondary layer. This is similar to the organization and secretion of the shell of bivalve molluscs.
Note the ornamentation, or sculpture of the shell surface. In some species radial grooves and ridges extend from the posterior end outward to the anterior margin. (Some genera do not have radial ridges). There are also concentric growth ridges that mark periods of interrupted growth.
If you are studying the shell of a whole animal, you will have to interrupt your study while you examine its soft parts. When you have completed your study of the soft anatomy, remove the tissues from the shell and return to this section to continue your examination of the shell. If you are using a cleaned, empty shell, complete your study now.
A complex hinge with hinge teeth is present at the posterior end of the shell and is where the two valves articulate with each other. The hinge teeth maintain the proper alignment between the valves as they open and close. Put the two valves together so the hinge teeth articulate posteriorly and note the closeness of the fit between the two valves. The hinge of the ventral valve bears a pair of lateral teeth that articulate with deep sockets in the hinge of the dorsal valve. The ventral valve, of course, bears the aperture for the pedicle.
The dorsal valve supports the lophophore. It bears a pair of slender, delicate crura that extend from the hinge into the body of the brachiopod (Fig 25-8E). In life these are the points of attachment of the skeleton, or brachidium, of the lophophore. The crura are delicate and may not be present in cleaned shells. The brachidium is also calcareous but is exceedingly delicate and is never present in routinely cleaned shells. There may be a median, longitudinal ridge inside the dorsal valve. This is the septum to which the brachidium attaches. (The septum is absent is some taxa).
The posterior, medial edge of the hinge of the dorsal valve is extended to provide a site for the insertion of the abductor muscles (= diductor muscles), which open the shell (Fig 25-8C, 25-9D). This extension is the cardinal process. Place the two valves together and articulate them at the hinge. Open and shut the valve, noting that the dorsal valve rotates on a transverse axis that passes through the two hinge teeth. The teeth form a fulcrum. Note that the cardinal process is posterior to the fulcrum. Imagine a muscle (the abductor muscle) running from the cardinal process to the center of the ventral valve and think about the effect it would have when it contracts.
Other muscles (the adductor muscles) extend from the ventral valve to a position on the dorsal valve anterior to the hinge (Fig 25-8C, 25-9D). Visualize the position of this muscle and the effect its contractions would have on the relative positions of the two valves. The muscles will be discussed more fully in a later section.
Body
Place a preserved specimen in a small dissecting pan of tapwater. Locate the dorsal and ventral valves of the shell. A fleshy, tough pedicle can be seen emerging from the aperture in the ventral valve (Fig 1, 25-8C). The pedicle is an attachment organ by which the brachiopod anchors itself to hard substrata, usually rocks (Fig 25-9C). In life the pedicle usually holds the shell with the larger ventral valve uppermost. The pedicle is often cut or damaged when the animals are collected.
Figure 1. Ventral view of Terebratella. The ventral valve and ventral mantle lobe have been removed. The ventral surface of the body, lophophore, and muscles are thus revealed. Brach30La.gif
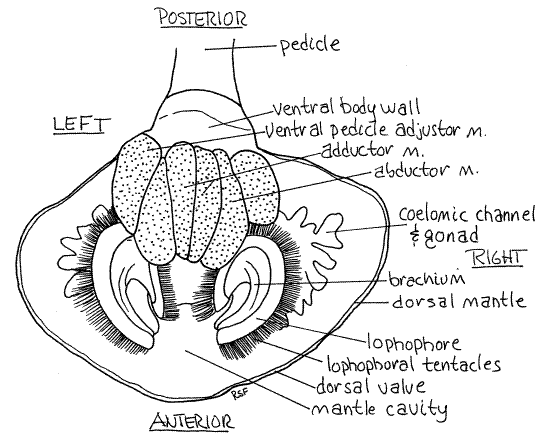
Like that of a bivalve mollusc, the brachiopod body wall is known as the mantle which forms two thin folds of the body wall called the mantle lobes. Unlike those of any bivalve, however, the mantle lobes of brachiopods are dorsal and ventral and they are folds of the anterior, rather than dorsal, body wall. The two mantle lobes enclose mantle cavity and secrete the shell (Fig 3). They lie against the inner surface of the valves.
" Open the shell to expose the soft parts of the animal. First slip the blade of a scalpel between the anterior margins of the two valves and twist the blade to pry the valves apart. Hold them apart with your fingers and find the thin mantle lobe lying tightly against the inner surface of each valve. Slip the scalpel blade between the ventral mantle lobe and the ventral valve and use it to separate them from each other. Do not cut the mantle, rather use the scalpel to lift it from the valve.
Work the mantle free from the ventral valve beginning at the anterior edge and proceeding posteriorly. In the posterior third of the valve you will encounter the muscles that hold the two valves together and open and close them. Use the scalpel to scrape these muscles from their insertions on the valves. Do not cut the muscles, rather scrape them free of the valve with the scalpel. When the muscles are freed, remove the ventral valve. Set it aside intact. The animal will remain attached to the smaller dorsal valve. Do not try to pull the animal away from the dorsal valve. Conduct the dissection from the ventral surface with the animal attached to the dorsal valve.
Look at the exposed ventral mantle lobe (Fig 25-10). Lift its anterior edge and confirm that it is a thin sheet of tissue. Be aware, however, that as a fold it is a double layer of the body wall. Between the two walls are extensions of the trunk coelom (= metacoel). Coelomic channels extend from the coelom into the mantle and the gonads are located in their mesothelial walls. The coelomic channels and gonads are easily seen (Fig 1, 3). The presence of gametes in the gonads makes them much thicker than the remainder of the mantle lobe.
Note that the mantle lobes are attached to the body posteriorly but are free and unattached anteriorly. The inner surface of the mantle is ciliated. There is a similar dorsal mantle lobe inside the dorsal valve (Fig 1). The space between the dorsal and ventral mantle lobes is the mantle cavity. In life it contains seawater. The two large curved arms, or brachia, of the lophophore occupy most of the space in the mantle cavity (Fig 1, 25-10). The lophophore will be studied in more detail later.
" The valve musculature forms a large mass at the posterior end of the ventral surface. Use your fine scissors to remove the ventral mantle lobe by cutting around the lateral and anterior borders of this muscle mass.
The thin membranous, white, ventral body wall can be seen on the ventral surface around the base of the pedicle. The lateral body walls are visible as more or less vertical sheets dorsal to the edges of the sides of the muscle mass. Avoid damaging the body wall.
Lophophore
The lophophore is composed of two coiled arms, or brachia, joined on the midline above the mouth (Fig 1, 3, 25-10). It bears a fringe of ciliated tentacles. Look at the ventral surface and relocate the origin of the adductor muscle. Just anterior to the adductor you will see the portion of the lophophore that crosses the midline and connects the right and left brachia. Follow the ridge of the lophophore to the right or left as far as it goes. You will end up back where you started. The ridge bears the row of hollow, ciliated tentacles that extends its entire length.
Paralleling the row of tentacles is a thin, elevated, flap of tissue, the brachial fold. The valley between the row of tentacles and the brachial fold is the brachial groove (Fig 2, 25-13B). It is a ciliated groove that transports food from the tentacles to the mouth. Look again at the transverse ridge of the lophophore where it crosses the midline in front of the adductor muscle. Push the tentacles and brachial fold away from each other to reveal the mouth, which is located on the midline. The brachial fold is anterior to the mouth, the row of tentacles is posterior to it.
The lophophore is supported by a skeletal element, the brachidium. The tortuously curved, calcareous brachidium more or less parallels the coils of the brachia and supports them. A U-shaped arm of the brachidium, with dorsal and ventral portions, supports each brachium. The brachidium is easily seen and felt (it is hard) at the base of the row of tentacles. It begins and ends with the two crura of the dorsal valve and consists of a dorsal loop and a ventral loop. Find the brachidium and trace it through its two loops.
Brachidium
> 1a. The internal lophophoral skeleton, or brachidium, can be exposed by treatment with hot concentrated potassium hydroxide. Do not do this with you dissection specimen unlesss you are finished with the soft anatomy. If you undertake this procedure be very careful with the potassium hydroxide pellets and solution. Both are very corrosive and can cause serious burns. Wear safety glasses and work in a fume hood.
Remove the ventral valve from a large, undissected specimen but leave the dorsal valve intact and attached to the animal. Do not use the same specimen you are dissecting as it will be destroyed. Handle this specimen very carefully throughout the procedure as the brachidium is very fragile. Place the specimen with the dorsal valve down in a 100 ml beaker. Cover the specimen to twice its depth with water and add enough potassium hydroxide pellets or flakes to make a concentrated solution. Place the beaker on a hotplate and heat the solution but do not boil it. The tissue will decompose leaving the brachidium, hopefully intact and still attached to the crura. Handle it very carefully and rinse it several times with fresh water. <
Figure 2. Dorsal surface of the anterior body of the inarticulate brachiopod Lingula with the brachia removed. Brach29L.gif
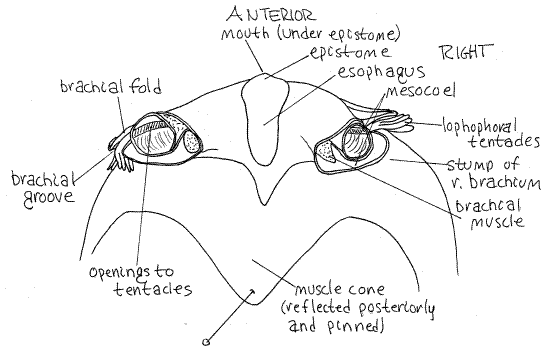
Muscles
The musculature that operates the valves is much simpler in articulates than in inarticulates. This is because the elaborate hinge of articulates prevents motion in all directions except up and down and only two sets of muscles are needed for this, one for opening (abduction) and one for closing (adduction). The valves of inarticulates, on the other hand, can move in almost any direction and must have muscles to cause or prevent motion in multiple directions. The resulting system is complex, as you well know if you have dissected an inarticulate such as Lingula or Glottidia.
In articulates there are only two pairs of valve muscles and another one or two pairs for the pedicle (Fig 3, 25-10B). The ends of these muscles have already been seen on the surface of the posterior part of the body.
The two adductor muscles pull the two valves together and close the gape (Fig 25-8C). Each originates on the ventral valve beside the ventral midline and runs straight across the coelom to insert by two heads on the dorsal valve. There are thus four points of insertion for the two muscles on the dorsal valve. At present you can see only the ventral ends (origins) where there is a total of only two heads (Fig 1). These touch each other on the midline of the ventral valve. The origin and insertions of the adductors are anterior to the fulcrum of the hinge. Contraction of the muscles pulls the two valves toward each other (adducts the valves).
The two abductor muscles (= diductor muscles) open the valves (Fig 3, 25-8C). They have no muscular equivalent in bivalve molluscs but their functional equivalent is the bivalve hinge ligament. They originate on the ventral valve lateral to and touching the origins of the adductors. They also have large origins but the belly is narrow. The origins are long narrow ovals lateral to the origins of the adductor muscles. The abductors narrow rapidly as they cross the body cavity to insert on the dorsal valve. They extend posterodorsally from the ventral valve to insert on the cardinal process at the posterior end of the dorsal valve. The origin is anterior to the fulcrum but the insertion is posterior to it. Consequently, contraction of these muscles pulls the posterior end of the dorsal valve down and causes its anterior end to move up, like a see-saw. It thus moves the valves apart and opens the gape.
A set of pedicle adjustor muscles moves the shell with respect to the pedicle (Fig 3, 25-10B). In Terebratella there are two pairs of these muscles. The two ventral pedicle adjustor muscles originate on the ventral valve lateral to and touching the origin of the abductor muscles (Fig 1, 3). They extend obliquely posteriorly medially and dorsally to insert on the pedicle.
It is the origins of these six muscles that make up the large muscle mass occupying the posterior part of the ventral surface of the animal. There is also a pair of dorsal pedicle adjustor muscles which you cannot see yet. They do not contribute to the ventral muscle mass.
Internal Anatomy
" With fine scissors open the body cavity (trunk coelom) by removing the thin body wall between the base of the pedicle and the ventral muscle mass (Fig 3, 25-10). The cavity lying dorsal to the body wall, which you have just opened, is the trunk coelom, or metacoel. You have opened its posterior ventral corner.
Look inside the posterior trunk coelom and visually trace the muscles as far as you can without further cutting. Do not alter any tissues in the coelom. Note that the adductors run vertically across the coelom but the abductors run posteriorly and dorsally, and the ventral pedicle adjustors extend to the base of the pedicle.
" Using fine scissors carefully separate the ventral edge of the lateral body wall on the right side from the origin of the abductor and pedicle adjustor muscles. Be careful that you do not damage this wall, especially its dorsal portion. Separating it from the muscles gives you a better look into the ventral, posterior part of the body coelom. It is possible that this wall is already damaged or missing. To improve your view further, remove the right abductor and right ventral pedicle adjustor muscles by cutting across their narrow bellies. Do not remove the adductor muscle.
Figure 3. Sagittal section of the articulate brachiopod Terebratulina. Redrawn from Williams and Rowell, 1965). Brach33L.gif
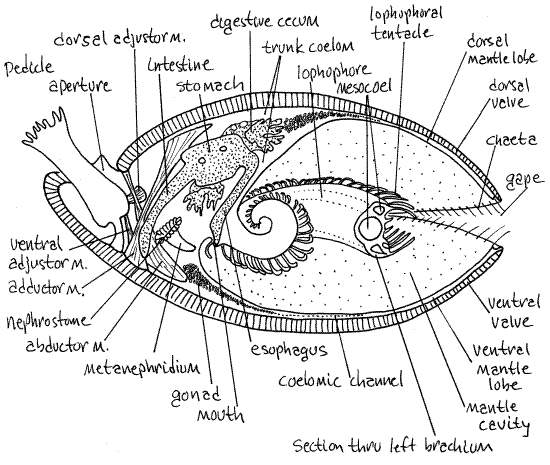
A large bulging metanephridium is visible on the inside surface of the lateral body wall (Fig 3, 25-10A). It is a ciliated funnel extending from the trunk coelom to the mantle cavity. The coelomicl opening of the funnel is the nephrostome. It has a folded border and faces posteriorly. A wide tubule runs from the nephrostome anteriorly along the lateral body wall to empty via a nephridiopore on the anterior body wall ventral to the lophophore. The nephridiopore is difficult to find. This metanephridium is one of a pair. Some articulate brachiopods have two pairs but Terebratella has only one.
Pull the ventral end of the combined adductor muscles anteriorly so you can see the posterior edge of the muscle as it crosses the coelomic cavity. The large vertical tube attached to the posterior edge of the muscle by a mesentery is the intestine (Fig 3, 25-10). In articulates the intestine ends blindly and does not open to the exterior, there being no anus. The medial edges of the nephrostomes are connected to the intestine by mesenteries.
Note the thin, delicate, calcareous projections from the hinge region of the dorsal valve. These are the two crura (singular = crus) (Fig 25-8E). They extend to the base of the lophophore and are the points of attachment of a calcified band, the brachidium, that supports the lophophore. The brachidium is a continuous loop extending from one crus to the other following the curves of the lophophore.
" It is not practical to continue the dissection further from the ventral side. Use your scalpel to free the dorsal mantle lobe from the dorsal valve. Separate the insertions of the adductor and abductor muscles from the dorsal valve and detach the lophophore from the crura. The connections between the brachidium and the crura are very delicate and usually break of themselves long before you deliberately break them. Remove the animal from the dorsal valve and orient it in the pan with the dorsal side up, facing you.
Look briefly at the dorsal mantle lobe (Fig 3, 25-10) to confirm its resemblance to the ventral lobe, including the presence of coelomic channels filled with gametes. Note that the muscle mass is surrounded by the dorsal body wall, inside of which is the trunk coelom. The muscle mass consists entirely of the four heads of the two adductor muscles. The dorsal ends of the abductors insert far posteriorly on the cardinal process and do not contribute to this mass. The dorsal pedicle adjustor muscles, which you are seeing for the first time, are small and run from the hinge region of the dorsal valve to the ventrolateral surface of the pedicle (Fig 3).
" Trim away the dorsal mantle lobe and remove the dorsal body wall. Find once again the intestine but in dorsal view this time. Trace it anteriorly and dorsally to the point where it exits the much wider stomach (Fig 3, 25-10B). The stomach is connected to the median mouth by the esophagus. The heart is a small spherical organ on the dorsal edge of the stomach at the origin of the intestine (Fig 25-10A). An aorta can be seen running over the dorsal midline of the stomach. The densely papillated organs in the vicinity of the stomach are the digestive ceca (Fig 3, 25-10A,B). The ceca are diverticula of the stomach and connect with it via ducts. Return to the stomach and open it with a midventral incision. Note the internal folds of its walls. Find its posterior opening into the intestine and its small anterior opening to the esophagus. Trace the esophagus anteriorly and ventrally to the mouth by opening its lumen with fine scissors.
Shell
If you have not studied the interior of the shell, do so now. Remove the soft parts from the interior of the shell. Return to the section on the shell earlier in this exercise and find the features described there.
References
Abbott DP . 1987. Observing Marine Invertebrates. Stanford University Press, Palo Alto. 380p. (Terebratalia and Laqueus californianus).
Bullough WS. 1958. Practical Invertebrate Anatomy. MacMillan, London. 783p.
Hyman LH. 1959. The Invertebrates 5. Smaller Coelomate Groups. McGraw-Hill, New York. 783p.
Pierce M.E. 1950. Terebratulina septentrionalis or Terebratella in F. A. Brown (ed) Selected Invertebrate Types. Wiley, New York. pp 357-359.
Rudwick MJS. 1970. Living and Fossil Brachiopods. Hutchinson University Library, London. 199p.
Ruppert EE, Fox RS, Barnes RB. 2004. Invertebrate Zoology, A functional evolutionary approach, 7 th ed. Brooks Cole Thomson, Belmont CA. 963 pp.
Williams A, Rowell AJ. 1965. Brachiopod anatomy, pp. H6-H57 in Moore RC (ed), Treatise on invertebrate paleontology, Part H, Brachiopoda, vol 1. Geological Society of America and University of Kansas Press, Lawrence, KS.
Supplies
Dissecting microscope
Small dissecting pan
Articulate brachiopod
Dissecting set with microdissecting tools
Isotonic magnesium chloride for living specimens
Empty, cleaned brachiopod shell
Bleach
For brachidium preparation: undissected specimens, potassium hydroxide pellets, 100-ml beakers, hot plate, safety glasses