Invertebrate Anatomy OnLine
Tectura testudinalis ©
Limpet
3jul2006
Copyright 2003 by
Richard Fox
Lander University
Preface
This is one of many exercises available from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises can be accessed by clicking on the links to the left. A glossary and chapters on supplies and laboratory techniques are also available. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Mollusca P, Eumollusca, Conchifera, Ganglioneura, Rhacopoda, Gastropoda C, Prosobranchia sC, Patellogastropoda O, Patellacea SF Acmaeidae F (Fig 12-125)
Mollusca P
Mollusca, the second largest metazoan taxon, consists of Aplacophora, Polyplacophora, Monoplacophora, Gastropoda, Cephalopoda, Bivalvia, and Scaphopoda. The typical mollusc has a calcareous shell, muscular foot, head with mouth and sense organs, and a visceral mass containing most of the gut, the heart, gonads, and kidney. Dorsally the body wall is the mantle and a fold of this body wall forms and encloses that all important molluscan chamber, the mantle cavity. The mantle cavity is filled with water or air and in it are located the gill(s), anus, nephridiopore(s) and gonopore(s). The coelom is reduced to small spaces including the pericardial cavity containing the heart and the gonocoel containing the gonad.
The well-developed hemal system consists of the heart and vessels leading to a spacious hemocoel in which most of the viscera are located. The kidneys are large metanephridia. The central nervous system is cephalized and tetraneurous. There is a tendency to concentrate ganglia in the circumenteric nerve ring from which arise four major longitudinal nerve cords.
Molluscs may be either gonochoric or hermaphroditic. Spiral cleavage produces a veliger larva in many taxa unless it is suppressed in favor of direct development or another larva. Molluscs arose in the sea and most remain there but molluscs have also colonized freshwater and terrestrial habitats.
Eumollusca
Eumollusca is the sister taxon of Aplacophora and includes all molluscs other than aplacophorans. The eumolluscan gut has digestive ceca which are lacking in aplacophorans, the gut is coiled, and a complex radular musculature is present.
Conchifera
Conchifera, the sister taxon of Polyplacophora includes all Recent molluscs other than aplacophorans and chitons. The conchiferan shell consists of an outer proteinaceous periostracum underlain by calcareous layers and is a single piece (although in some it may appear to be divided into two valves). The mantle margins are divided into three folds.
Ganglioneura
Most Recent molluscs are ganglioneurans, only the small taxa Aplacophora, Polyplacophora, and Monoplacophora are excluded. Neuron cell bodies are localized in ganglia.
Rhacopoda
The mantle cavity is posterior in the ancestor although it may be secondarily moved anterior by torsion. This taxon includes gastropods and cephalopods.
Gastropoda C
Gastropoda is the largest molluscan taxon and is the sister group of Cephalopoda. Gastropods are united by descent from a torted ancestor although many exhibit various degrees of detorsion. Many are coiled and asymmetrical but the ancestor was probably symmetrical. Gastropods are relatively unspecialized molluscs known collectively as snails. The univalve shell, present in the ancestral gastropod and in most Recent species, is reduced or lost in many representatives. The flat creeping foot was inherited from their eumolluscan ancestors but gastropods have developed a distinct head with an abundance of sophisticated sense organs. The originally posterior mantle cavity has become anterior as a consequence of torsion, although detorsion has reversed this condition in many. Gastropods were originally gonochoric and most remain so but many derived taxa are hermaphroditic. Most are marine but many taxa have invaded freshwater and the only terrestrial molluscs are gastropods. Most have a single gill, atrium, and nephridium but the most primitive representatives have two of each. Only one gonad, the right, is present. The ancestor probably had an operculum. The nervous system is streptoneurous (twisted by torsion).
Prosobranchia sC
Prosobranchia was once one of three great gastropod subclasses but it is no longer considered to be a monophyletic taxon although the concept continues to be used as a pedagogical convenience. Prosobranchs are the gastropods most like the ancestral snails. They are torted and most have a shell and are coiled and asymmetrical. The mantle cavity is anterior. Most are gonochoric and most have an operculum. Most have only one gill in the mantle cavity but some primitive taxa have two. The right atrium is lost in most. Prosobranchs are specialized for life in marine benthic habitats although representatives are also found in freshwater and on land.
Patellogastropoda O
Patellogastropods are limpets with symmetrical, non-coiled, conical shells. A recent interpretation proposes that the ancestral gastropod was perhaps a patellogastropod. This hypothesis is based on evidence that patellogastropod symmetry is primary and not a secondary development.
Acmaeidae F
The true limpets belong to Acmaeidae. They are limpet-shaped, with a large body whorl and no spire, but do not have the apical aperture characteristic of keyhole limpets. Water enters and leaves the anterior mantle cavity, entering on the left and exiting on the right. They, like most prosobranchs, have lost the right gill and atrium. Unlike most other prosobranchs, but like the keyhole limpets, they have both nephridia. The keyhole limpets, on the other hand, retain two gills, two atria, as well as two nephridia.
Laboratory Specimens
This exercise is written specifically for Tectura testudinalis (= Acmaea testudinalis). Tectura is a common intertidal and high subtidal species of the northeastern rocky coast of North America from the Arctic to New York. Any of the true limpets, of which there are many species on the west coast, can also be used.
The dissection should be conducted on the stage of a dissecting microscope in a small dissecting pan of isotonic magnesium chloride (if living) or tapwater (if preserved). Begin with the animal in sea water (if living) and then transfer it to magnesium chloride prior to the dissection.
External Anatomy
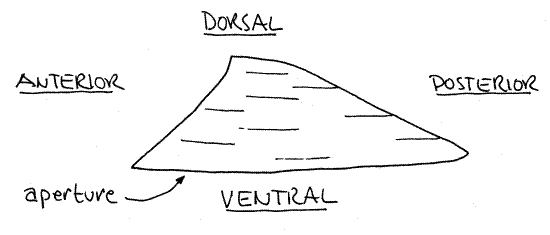
Make an initial study of the external anatomy with the shell intact. The term limpet refers to a distinctive body organization that has evolved several times in gastropods. It consists of a low dome-shaped shell derived from the body whorl of the ancestral coiled shell (Fig 1). The limpet shell is not coiled and has no spire. The shell is low and presents a reduced and streamlined profile to water currents. The foot is broad a flat and well adapted for clinging tenaciously to hard substrata in rough water. The similarities between the different limpet groups are due to convergence.
Shell
True limpets are not coiled but the body is torted. The shell is an oval cup that covers the dorsal surface of the animal. From the side it looks like a low hill or volcano but there is no opening at the apex as there is in the keyhole limpets (Fig 1). The slope of the hill is steepest anteriorly and the summit is displaced slightly anteriorly.
Ventral Features
Turn the animal on its back and examine the ventral surface. The large oval opening in the base of the shell is the aperture through which the foot is extended. The large, oval foot is used to clamp the animal to rocky substrata and for locomotion over the substratum. The visceral mass sits dorsal to the foot and is covered by the shell. The head is anterior to the foot and visceral mass.
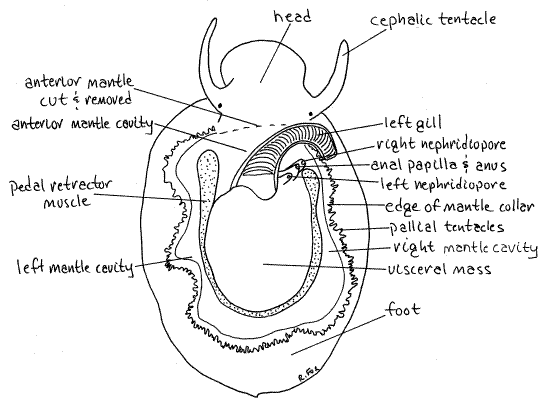
The mantle is the dorsal body wall and is large, thin sheet of tissue covering the dorsal surface under the shell (Fig 12-24). Most of it is now covered by the shell. Laterally the mantle is folded outward to form a collar, or skirt, extending away from the sides of the visceral mass and making a complete oval around the animal. The mantle skirt lies against the margin of the aperture of the shell and secretes the outer two layers of the shell (periostracum and prismatic layers). It is thickened here and bears short sensory tentacles (Fig12-24).
Figure 1. Side view of the shell of Tectura testudinalis. Gastrop165L.gif
Two lateral pallial grooves run longitudinally between the mantle skirt and the foot; one on the right and one on the left. Since these grooves are formed by the overhanging mantle skirt, they are in fact mantle cavities. To avoid confusing them with the mantle cavity proper they are referred to as pallial grooves.
The head extends anteriorly from the anterior wall of the visceral mass (Fig 2, 12-24). It bears two long cephalic tentacles and the mouth is situated ventrally at its anterior end. A tiny black eye can be seen at the base of each tentacle. The buccal mass is a vague reddish (in life) area inside the head.
Above the head a large anterior fold of the mantle covers a deep recess, the mantle cavity, at the dorsal anterior end of the visceral mass (Fig 2, 12-24). A single long feathery, bipectinate gill may be visible extending from the right side of the mantle cavity (Fig 2, 12-24).
If your specimen is living, replace the seawater in the dish with isotonic magnesium chloride. The animal will relax in about 5 minutes. If it is preserved, leave it in tapwater.
When placed in magnesium chloride, the snail may initially extend the foot far away from the shell. If it does, take advantage of the opportunity to see the pedal retractor muscle which runs vertically from the foot to the shell on both sides of the animal. It can be seen through the integument on the sides of the visceral mass. This muscle is formed of the fused right and left columellar muscles. Together they make a horseshoe, open anteriorly, around the body (Fig 2).
The ventilating current approaches the body from the left side of the head (Fig 12-24A). Some of it passes posteriorly in the left pallial groove and some of it enters the left side of the mantle cavity. In the mantle cavity the stream of water passes over the gill and between its filaments, over the openings of the kidneys and intestine, and then exits the right side. Here it enters the right pallial groove and is moved posteriorly along the right side of the animal. At the posterior end of the body the currents from the left and right pallial grooves join and move away from the animal. These currents are generated by the ciliated epithelium of the mantle and gill.
>1a. If you have a living specimen, use a suspension of carmine particles in seawater to visualize these currents. <
Mantle Cavity
" Insert the blade of a scalpel between the mantle and shell and use it to separate the two from each other. Use a careful scraping motion of the scalpel to separate the pedal retractor muscle from its origin on the inner surface of the shell and then remove the snail from the shell. It is held to the shell by the muscle.
Make a quick study of the major anatomical features disclosed by removing the shell before turning your attention to the mantle cavity and its contents. The dorsal body wall and the inner epithelium of the mantle cavity are a brilliant emerald green (in life).
The dorsal body wall is thin and transparent and some of the viscera can be seen through it. Some of the more conspicuous structures should be found at this time as they make convenient landmarks. The heart is in a white, triangular, pericardial cavity on the left side, and can be seen beating if it has not been in magnesium chloride too long. You may want to return the specimen to seawater so the heart can resume beating. The origin of the pedal retractor muscle makes a bright white oval encircling three sides of the visceral mass (Fig 2). The posterior 2/3 of the visceral mass is occupied by the brown digestive cecum composed of masses of small branched tubes (Fig 3).
" With fine scissors, carefully remove the roof of the mantle cavity so you can see the interior of the cavity. Do not penetrate the posterior wall of the mantle cavity yet.
Look into the mantle cavity and find the left gill. It attaches on the left side and extends obliquely across the body to emerge on the right. It is the left gill. The right one is absent as it is in advanced vetigastropods and all meso- and neogastropods.
Find the flat central axis of the gill from which arise two rows of semicircular filaments. One row lies on the left, the other on the right, of the axis. The gill, unlike those of meso- and neogastropods but like those of vetigastropods, is bipectinate. There is no hypobranchial gland. The obscure osphradium is located on the left side of the mantle cavity at the base of the gill axis.
On the posterior wall of the mantle cavity, to the right of the base of the gill, are three short tubes, stacked on top of each other. The ventralmost of the three is the right nephridial papilla (Fig 2). The nephridiopore of the right kidney opens at its tip. The middle and largest tube is the anal papilla and it bears the anus at its tip (Fig 2). Dorsalmost, and furthest to the left is the left nephridial papilla with the pore of the left kidney at its tip (Fig 2). Note that there are two kidneys in Tectura even though there is only one atrium and one gill. The left kidney is small but the right is very large.
The gonad shares the duct of the right kidney and gametes exit via the right nephridiopore into the right side of the mantle cavity where they are swept into the sea by the exiting respiratory current. In the higher gastropods the left kidney is retained as the sole functional kidney but the duct of the right persists as part of the gonoduct. The rest of the right kidney is lost.
Internal Anatomy
Structures will be considered in the order in which they appear in dorsal dissection.
Figure 2. Dorsal view of a true limpet, Tectura testudinalis, from Pemaquid Point, Maine. The shell and anterior mantle have been removed. Gastrop166L.gif
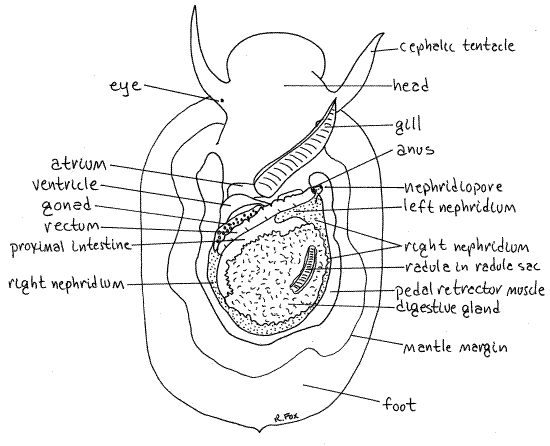
" The visceral mass contains most of the internal organs and occupies a concavity atop the foot (Fig 2). Use your fine scissors and fine forceps to remove the transparent dorsal mantle, i.e. body wall, from the surface of the visceral mass. Do this very carefully as many of the organs under it adhere to it. The lamellar layer of the shell is secreted by the outer surface of this part of the mantle. The cavity exposed by removing the dorsal mantle is the hemocoel.
The most obvious structure in the visceral mass is the finely branched, brown digestive ceca which occupies most of the posterior right 2/3 of the hemocoel (Fig 3). The rectum is a conspicuous white tube running obliquely across the anterior left corner of the hemocoel. It runs to the anal papilla. A short section of the wide, brown, proximal end of the intestine lies between the rectum and the digestive gland.
A section of the exceedingly long radula may be seen looking like a piece of a snakeskin on the surface of the digestive gland (Fig 3).
The heart occupies the extreme anterior left corner of the hemocoel. It consists of a muscular, contractile ventricle and a single atrium (Fig 3). Both chambers have colorless, transparent walls. The heart is located at the posterior end of the gill and is enclosed in a transparent membranous pericardium.
The long axis of the ventricle is transverse and an aorta exits each end of it. The atrium is anterior to the ventricle and drains oxygenated blood from the gill which lies to the right of it.
The large, black right nephridium is conspicuous on the right and posterior left sides of the visceral mass (Fig 3). It can be seen around the margins of the digestive gland. The left nephridium is inconspicuous and will not be seen. This resembles the relationship between the kidneys of the keyhole limpets.
Figure 3. Dorsal view of a true limpet with the dorsal mantle (body wall) removed to reveal the viscera and hemocoel. Gastrop167L.gif
Reproductive System
There is controversy regarding the sexuality of acmaeids. They are variously claimed to be either gonochoric or protandric hermaphrodites. The evidence for hermaphroditism is based on correlations of sex with body size. The same arguments have been made for the keyhole limpet, Diodora.
The single gonad is very large but only a small portion of it can usually be seen in dorsal view without further dissection (Fig 3). It is a large pink mass lying ventral to the digestive gland and gut. A small edge of it can sometimes be seen between the ventricle and the rectum. The gonad empties into the right kidney and the gametes exit via the right nephridiopore. Fertilization is external in this species.
Digestive System
" The mouth is located on the ventral surface of the head. With your fine scissors cut through the surface of the head to open the anterior gut.
The mouth opens into a small, vertical buccal cavity. The esophagus (or pharynx) extends posteriorly through the head and neck beginning at the top of the buccal cavity.
A very long pouch off the floor of the buccal cavity houses the radula. This is the radular sac and it extends, ventral to the gut, posteriorly into the digestive cecum where it loops dorsally and anteriorly. It is 1.5 times the length of the body as is the radula it contains.
" Open the buccal cavity and esophagus posteriorly, cutting through other structures as necessary.
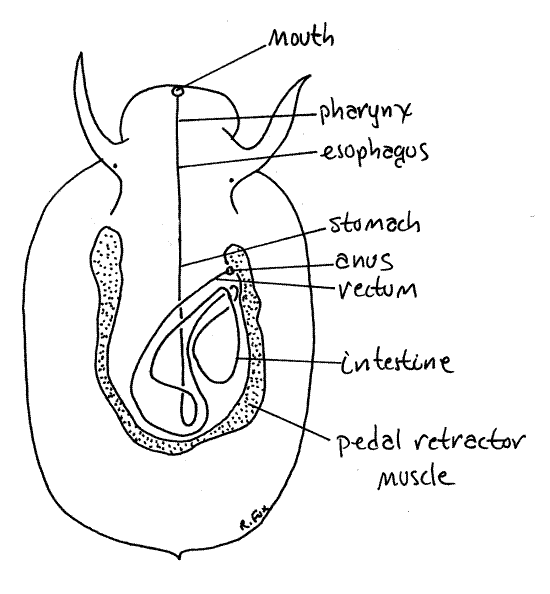
The esophagus leads from the buccal cavity to the long, thick-walled, yellowish-cream stomach lying on the midline (Fig 4). Open the stomach. The stomach has no gastric cecum such as is typical of many gastropods. A sphincter at the posterior end of the stomach separates it from the anterior end of the very long intestine.
The first, or proximal, portion of the intestine is wide and thin walled. A short section of it was seen earlier crossing obliquely over the surface of the digestive gland. The intestine makes several loops through the visceral mass and eventually becomes the rectum, which crosses the mantle cavity to empty via the anus (Fig 4).
The feces of true limpets are released into the mantle cavity where fouling of the gill and head is likely. To avoid this possibility the feces must be compacted into fecal pellets that will not disintegrate in the mantle cavity. A long intestine is required to accomplish this compaction.
Figure 4. Simplified dorsal view of Tectura. The labeled line is a diagrammatic representation of the route taken by the gut through the visceral mass. Gastrop168L.gif
" Remove the radula from the posterior part of the radular sac but leave it attached anteriorly. Stretch it out behind the snail and compare its length to that of the body.
>1b. Remove a short piece of the anterior radula and make a wetmount with it. Examine it with the compound microscope. <
" Cut around the lateral and posterior margins of the visceral mass and lift the posterior edge of the mass so you can see beneath it. This will give you a better idea of the extent of the gonad and kidney and will reveal some of the loops of the gut.
References
Abbott DP. 1987. Observing Marine Invertebrates. Stanford U. Press, Palo Alto. 380p.
Abbott D, et al. (eds) 1968. The biology of Acmaea. Veliger 11 (Suppl). (a collection of 11 undergraduate research papers on Acmaea).
Fretter V, Graham A. 1994. British Prosobranch Molluscs, 2 nd ed. Ray Society, London. 820 pp.
Ruppert EE, Fox RS, Barnes RB. 2004. Invertebrate Zoology, A functional evolutionary approach, 7 th ed. Brooks Cole Thomson, Belmont CA. 963 pp.
Walker CG. 1968. Studies on the jaw, digestive system, and coelomic derivatives in representatives of the genus Acmaea. Veliger 11(suppl):88-97.
Supplies
Dissecting microscope
Dissecting set with microdissecting tools
Living or preserved true limpet
Small dissecting pan (sardine can)
Isotonic magnesium chloride for living specimens