Invertebrate Anatomy OnLine
Taenia pisiformis ©
Dog Tapeworm
29may2007
Copyright 2001 by
Richard Fox
Lander University
Preface
This is one of many exercises available from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises can be accessed by clicking on the links on the left. A glossary and chapters on supplies and laboratory techniques are also available. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Bilateria, Protostomia, Platyhelminthes P, Neodermata, Cercomorpha, Cestoda C, Eucestoda sC, Cyclophyllidea O, Taeniidae F, (Fig 10-48)
Platyhelminthes P
Flatworms, or platyhelminths, are bilaterally symmetrical metazoans with three tissue layers. Unlike most triploblastic animals, they are compact and have no coelom (body cavity) surrounding the viscera and no hemal system. The gut, if present, has a single opening to the exterior. An anterior brain with associated concentration of sense organs is present, as expected of bilaterians. Flatworms are complex animals with elaborate hermaphroditic reproductive systems. Fertilization is internal with copulation. They may be free-living or parasitic.
Neodermata
Neodermata includes the parasitic flatworms, most of which are flukes or tapeworms. Most parasitic flatworms are endoparasites with complex life cycles requiring multiple hosts including a definitive host inhabited by the adult worm and one or more intermediate hosts inhabited by juvenile stages of the worm. The defining neoderm characteristic is the neodermis, or tegument. Neodermis is an epidermis specialized for living in a potentially hostile environment from which it must absorb food but reject toxins. The neodermis is a syncytium with its cell nuclei submerged below the basal lamina (Fig 10-33*, 10-36).
Cercomeromorpha
Cercomeromorphans have anchoring hooks on the posterior end, the cercomer, of the ciliated oncosphere larva. Such hooks are lacking in the sister taxon, Trematoda.
Cestoda
Cestodes, or tapeworms, are highly specialized internal parasites and with few exceptions the adults inhabit the intestine of a vertebrate. The life cycle almost always includes an intermediate host inhabited by a larval stage. Either invertebrates or vertebrates may be intermediate hosts. The intermediate host is usually the prey of the definitive host. The definitive host is a vertebrate.
The cestode body is composed of an anterior scolex, with which the worm attaches to its host, and a long posterior strobila which is divided into numerous segment-like units known as proglottids (Fig 10-47*). Most organ systems are reduced or absent except for the reproductive system, which is well developed. There is no gut and metabolism is anaerobic utilizing small organic monomers absorbed across the body surface. The syncytial neodermis bears microvilli to facilitate this absorption (Fig 10-45).
Cestodes are compact (acoelomate) worms whose interior is filled with unspecialized, mesenchymal connective tissue known as parenchyma, as it is in other platyhelminths. Excretion and osmoregulation are accomplished via protonephridia which empty into two pairs of excretory canals draining to the posterior end of the strobila. The nervous system includes anterior nerve rings, lateral longitudinal nerve cords, and transverse commissures. Cestodes are hermaphroditic and may utilize either self- or cross-fertilization.
Laboratory Specimens
Commercially prepared slides of Taenia pisiformis (= T. serrata), the dog tapeworm, are frequently used in the laboratory as examples of tapeworm anatomy. Other species can also be used but differ in some respects. The adults of Taenia pisiformis occur in dogs, cats, and other carnivores and the intermediate host is a rabbit. Use the compound microscope to study wholemount slides of the scolex, a mature proglottid, and a gravid proglottid.
The staining quality of commercial slides is variable. It is hard to see structures on dark heavily-stained, pale weakly-stained, or poorly fixed slides. You may need to look at several slides to see some structures and there may be other structures you simply cannot find.
Anatomy
Scolex
The tapeworm body consists of an anterior, head-like scolex and the trunk, or strobila, consisting of a linear series of segments, or proglottids. Look at the wholemount of a scolex with the scanning lens (40X) of the compound microscope (Fig 1, 10-44A). The scolex is wider than the strobila to which it joined by a narrow neck.
The scolex attaches the worm to the gut wall of the host. For this purpose it has a retractable rostellum armed with two rings of rostellar hooks. Examine the hooks with 100X power. Just posterior to the rostellum is a ring of four suckers; two dorsolateral and two ventrolateral. Scolex morphology varies widely with taxon (Fig 10-44).
Two lateral nephridial canals may be visible on each side of the scolex. They connect with each other near the rostellum via a set of convoluted scoelcial nephridial canals and extend posteriorly through the strobila. A pair of nerve rings ispresent but will not be evident.
Figure 1. The scolex and anterior strobila of Taenia pisiformis viewed from the ventral surface. Tape22La.gif
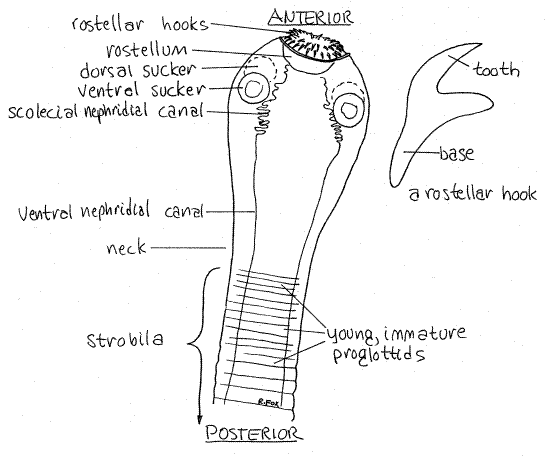
The anterior end of the strobila consists of the youngest immature proglottids. They are short and wide and do not yet contain differentiated cells or organs. New proglottids are produced by an area of mitotically active cells just behind the scolex. This is the opposite of segmented animals such as annelids and arthropods in which the segment-producing teloblast cells, and the youngest segments, are at the posterior end of the body.
Strobila
The strobila is long and wormlike, sometimes reaching 15 or more meters in length. (There are records of Taeniarhynchus saginatus (= Taenia saginata, the beef tapeworm), a human parasite, reaching lengths of more than 20 meters.) The strobila is strongly flattened dorsoventrally and is a linear series of hundreds or thousands of segment-like proglottids.
The strobila includes a series of young immature proglottids at its anterior end, then a region of sexually mature proglottids in the middle, and a length of gravid proglottids at the posterior end. The gravid proglottids contain viable embryonated “ eggs” ready to infect a new host should they get the opportunity. Proglottids increase in size posteriorly.
Each mature proglottid contains its own complete hermaphroditic reproductive system as well as its share of the common excretory and nervous systems. There is no digestive system in cestodes and predigested food monomers are absorbed across the neodermis. The reproductive systems of the youngest, anteriormost proglottids are not yet formed but more posterior proglottids have increasingly better-developed reproductive systems. Most tapeworms are protandric with the male system of each proglottid maturing first.
Examine a mature proglottid using 40X of the compound microscope (Fig 2, 10-47). Your slide probably contains several proglottids, some of which may be better than others. You will probably want to make use of them all as you look for the best representation of each structure.
Find the anterior and posterior ends of the proglottids. In Taenia, the posterior end of each proglottid is wider than the anterior end of the next proglottid (Fig 2).
Figure 2. Dorsal view of a mature proglottid of Taenia pisiformis. Tape23La.gif
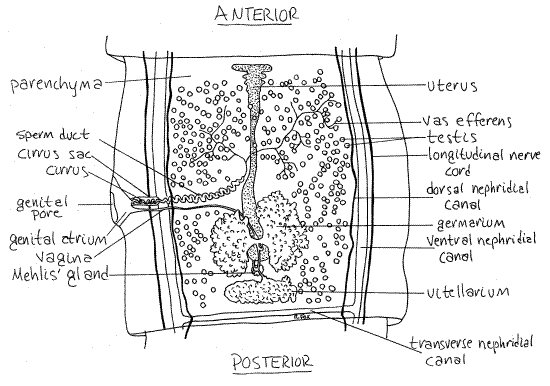
Body Wall
The body wall consists of a syncytial, microvilliated, absorptive neodermis (Fig 10-45), a basal lamina, and layers of circular and longitudinal muscles. The nuclei of the syncytium are submerged below the basal lamina and muscle layers into the parenchyma. Inside the body wall is connective tissue consisting of the mesenchymal parenchyma in which the reproductive, excretory, and nervous systems are embedded.
Digestive, Respiratory, and Fluid Transport Systems
Tapeworms have no mouth or gut. Organic molecules, mostly glucose, other sugars, and amino acids, are absorbed across the specialized, heavily microvillous neodermis and metabolism is largely anaerobic. There is no need for a gas exchange mechanism and there is none. Food and waste molecules diffuse to and from the body surface and there is no fluid transport system. The microvilli of the neodermis absorb food molecules from the environment.
Excretory System
The excretory system consists of numerous flame bulb protonephridia scattered throughout the parenchyma of each proglottid but they are not evident in these preparations. Individual protonephridia drain into an elaborate system of nephridial canals which ultimately open to the exterior at the posterior end of the last proglottid of the strobila. Dorsal and ventral lateral nephridial canals on each side of each proglottid extend the length of the worm. In addition to the two nephridial canals, each side possesses a longitudinal nerve cord which may be confused with the canals. The visibility of these structures depends on the quality of the staining and they are often indistinct.
The larger ventral nephridial canal is a pale, wide, longitudinal band lateral to the testes (Fig 2). The dorsal canal is much smaller in diameter and is located medial to the ventral canal, between it and the testis. In each proglottid the right and left ventral canals are connected by a transverse nephridial canal extending across the posterior edge of the proglottid just anterior to the junction with the next proglottid. The canals may be easier to see in immature proglottids.
Nervous System
The right and left lateral longitudinal nerve cords arise from nerve rings in the scolex and pass posteriorly in the sides of the proglottids (Fig 2). They are slender longitudinal lines lateral to ventral nephridial canals but are not visible in most preparations.
Reproductive System
Male
Most structures in the proglottid belong to the reproductive system. The male and female systems share a common genital pore (= gonopore) and genital atrium but are otherwise independent of each other. The common genital pore (Fig 2) is a large aperture on either the right or left side of the proglottid. It opens into a shallow, cuplike genital atrium. The male and female systems both open into the atrium via its own gonoduct.
Find the two ducts joining the medial border of the genital atrium. The anterior duct is the thicker and is the male gonoduct. The gonoduct is regionally specialized. The wide portion of the gonoduct attached to the atrium is the muscular cirrus sac. Inside the sac is the convoluted, eversible, tubular cirrus, which is the intromittent organ, or penis. During copulation the cirrus is extended from the genital pore and inserted into the genital atrium and vagina of another proglottid. On your slide it may be everted in some proglottids, in which case it looks like a slender worm extending from the genital pore.
The next region of the male gonoduct is the tubular sperm duct , also convoluted, which extends to the testes. Its entire length is not visible. The testes are numerous small spheres scattered throughout the parenchyma. Each is drained by a tiny tributary of the sperm duct, but these cannot be seen. There is no seminal vesicle and autosperm are stored in the coils of the sperm duct.
Figure 3. Details of the female reproductive system in dorsal view. Tape24La.gif
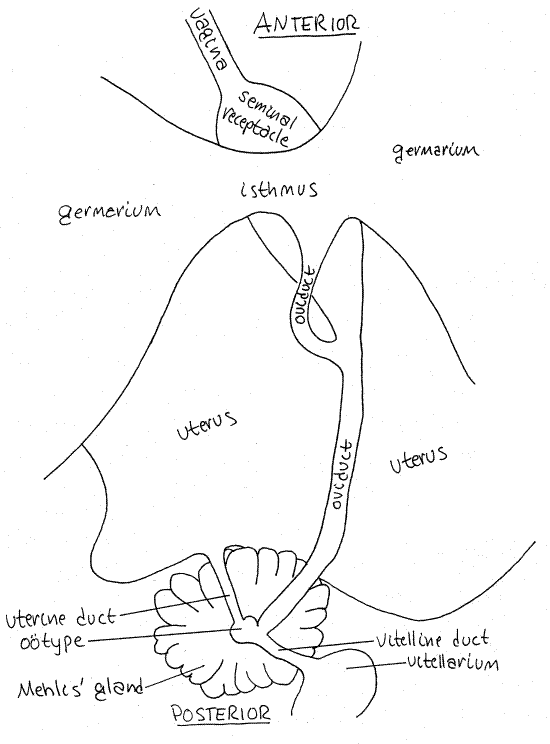
Female
The smaller and more posterior of the two ducts entering the genital atrium is the female gonoduct, which is also regionally specialized. The first region is the vagina. It receives the partner's cirrus and sperm during copulation. The vagina extends medially and posteriorly to the small seminal receptacle (Figs 2, 3, 10-47). This is a clear, unstained, oval chamber where allosperm received by the vagina are stored. It is usually easily visible. A short duct exits the posterior end of the seminal receptacle and joins the oviduct.
The germarium (= ovary) is divided into large right and left lobes lying on either side of the seminal receptacle. It is the site of oogenesis and produces large numbers of small, yolkless oocytes. The two lobes of the germarium are connected across the midline by a short, wide, transverse isthmus (Fig 3).
The follicles of the germarium open into small ducts which drain into the isthmus. The narrow oviduct arises from the isthmus and extends posteriorly for a short distance before receiving the duct from the seminal receptacle (Fig 3). The isthmus is usually easy to see but the oviduct is often obscured by the seminal receptacle and is harder to find. After receiving the duct from the seminal receptacle the oviduct continues posteriorly to the ootype. Fertilization occurs in the oviduct.
Yolk cells are produced by the single vitellarium at the posterior end of the proglottid. A short vitelline duct exits the vitellarium and extends anteriorly to join the oviduct at the ootype. Mehlis’s gland surrounds the ootype. The ootype, Mehlis's gland, and associated ducts cannot be seen in most slides.
A small uterine duct, usually not discernible, extends from the ootype to the uterus (Fig 2,3). Shelled eggs move from the ootype through the uterine duct into the uterus. Within the shell meiosis is completed, a zygote forms, and development proceeds to the oncosphere larval stage. The uterus is a blind sac with lateral branches in which embryonated “eggs” are stored. The size and visibility of the uterus vary with the maturity of the proglottid.
As the proglottid ages the accumulating “eggs” cause the uterus to become larger, darker, and more visible. They will eventually fill it, distending it so it occupies the entire proglottid. There is no opening of the uterus to the exterior and eggs are released by rupture of the proglottid.
Fertilization and Development
Tapeworms are hermaphroditic and commonly self-fertilize, a convenience for an animal that lives in a habitat where it may be the only member of its species. Cross-fertilization may occur if more than one worm is present in the host.
Fertilization of oocytes (from the germarium) by sperm (from the seminal receptacle) occurs in the oviduct (Fig 3). The fertilized oocyte (with meiosis still in progress) moves posteriorly in the oviduct to the ootype where it associates with a yolk cell from the vitellarium. The role of Mehlis's gland is unclear but it may secrete a thin, delicate membrane around the yolk cells but it does not secrete the eggshell as was formerly thought.
The eggshell is the product of a joint effort by the yolk cell and the developing embryo. Shelled embryonated “eggs”, with development in progress, move anteriorly into the uterus where they accumulate and are stored. In the uterus meiosis is completed, a zygote is formed, cleavage ensues, and development advances to the oncosphere larva stage. Mature, infective “eggs” in the gravid uterus contain these larvae.
Gravid Proglottid
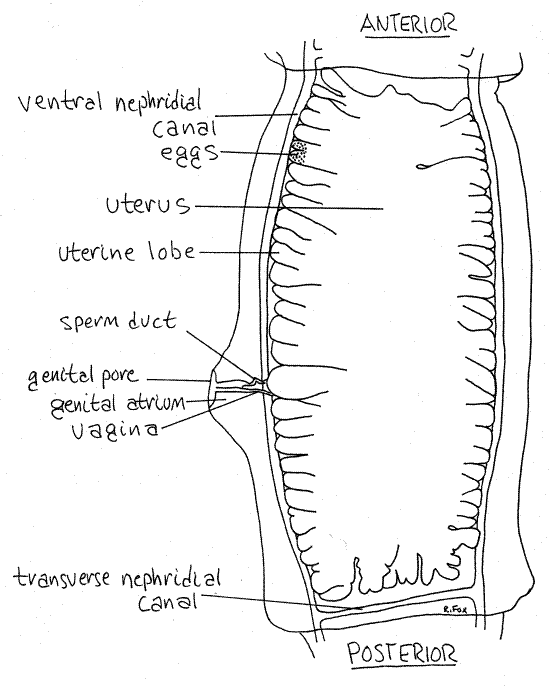
In the gravid proglottid the uterus is vastly expanded and packed with eggs (Fig 4, 10-47). Other parts of the reproductive system are degenerate and may no longer be identifiable but the nervous and excretory systems are present and functional, although not necessarily visible. The body wall musculature remains and gravid proglottids are mobile and very active.
Look at a slide of a gravid proglottid (Fig 4) and compare it with the mature proglottid. The uterus, filled with small spherical “eggs”, fills all available space. Its lateral uterine lobes are much larger now and so numerous they crowd against each other, filling most of the interior. They are bounded laterally by the ventral nephridial canals. The appearance and location of these canals is the same as in the mature proglottid although dorsal nephridial canals are usually not present in posterior proglottids.
The genital pore, genital atrium, sperm duct, and vagina are present but no longer functional. Remnants of other parts of the reproductive system may be visible also.
Figure 4. A gravid proglottid. Tape25La.gif
Life Cycle
The life cycle of the dog tapeworm, Taenia pisiformis, includes only one intermediate host although many tapeworms require two. In this species the intermediate host is a rabbit or hare and the definitive host is a carnivore, usually a dog.
In the intestine of the definitive host, “egg”-filled gravid proglottids break away from the posterior end of the strobila and are eliminated with the feces. Sometimes, after breaking away from the strobila, they migrate under their own power out the anus and onto the perianal region of the host. Pet owners may have noticed small, white, rice-shaped proglottids crawling about the hind ends of their pets. Proglottids may also be found scattered about on the furniture and floor after they have dried. Outside the body, the proglottids rupture, releasing thousands of infective "eggs", each containing an oncosphere larva. The oncosphere is a small mass of poorly differentiated cells with a pair of protonephridia, some muscles, and six hooks radiating from its body.
The eggs are ingested by the intermediate host (rabbit) and the oncosphere escapes into its gut. The oncosphere bores through the gut wall and penetrates an abdominal organ, usually the liver. Here it metamorphoses into a cysticercus larva, or bladderworm. The cysticercus is relatively large, swollen, and fluid-filled. It has an inverted scolex, with rostellum. If the rabbit is ingested by a carnivore, the cysticercus finds itself in the intestine of its definitive host. Here it everts its scolex, attaches to the gut wall, and begins production of its strobila to become an adult worm.
> a. Study slides of cysticercus larvae (bladderworms), at 100X, if they are available in your laboratory. Find the inverted scolex and rostellum and think about how it could invert to become functional scolex in the gut of the definitive host.
Human Tapeworms
Taeniidae includes two species that occur in humans. The beef tapeworm, Taeniarhynchus saginata (= Taenia saginata), is a human parasite for which cattle are the intermediate host. It is the most common large human tapeworm. Humans become infected by eating raw or poorly cooked beef but the worm causes little damage. Humans are also the definitive host of Taenia solium, the pork tapeworm (pigs are an intermediate host), but it is a more dangerous to humans than is T. saginatus. The danger comes, not from the adult worms in the intestine, rather from the cysticercus larva which, although they normally infest the intermediate host (a pig), can also develop in humans if onchospheres are ingested. The cysticerci, known also as bladderworms, create and inhabit large fibrous cysts in virtually any tissue of the body, including the brain where they are extremely dangerous. Adult tapeworms, on the other hand, usually do surprisingly little damage and appropriate very little of the host's resources.
*Hyphenated call-outs, such as this one, refer to figures in Ruppert, Fox, and Barnes (2004). Those without hyphenation refer to figures embedded in this exercise.
References
Brown FA (ed). 1950. Selected Invertebrate Types. Wiley, New York. 597p.
Coil WH. 1991. Platyhelminthes: Cestoidea, in Harrison FW, Bogitsh BJ (eds.). Microscopic Anatomy of Invertebrates vol. 3 Platyhelminthes and Nemertinea . Wiley-Liss, New York. 347p.
Hyman LH. 1951. The Invertebrates: Platyhelminthes and Rhynchocoela, vol. II. McGraw-Hill, New York. 550p.
Schmidt GO, Roberts LS . 1989. Foundations of Parasitology, 4 th ed. Times Mirror/Mosby, New York. 750p.
Ruppert EE, Fox RS, Barnes RB. 2004. Invertebrate Zoology, A functional evolutionary approach, 7 th ed. Brooks Cole Thomson, Belmont CA. 963 pp.
Supplies
Compound microscope
Wholemount slides of scolex, mature proglottid, gravid proglottid of Taenia pisiformis.
|
Item |
Source |
|
Taenia pisiformis wholemounts |
Triarch, Carolina, Ward’s |
|
cysticercus |
Carolina , Ward’s |