Invertebrate Anatomy OnLine
Sclerodactyla briareus ©
Sea Cucumber
with notes on Cucumaria
5jul2006
Copyright 2001 by
Richard Fox
Lander University
Preface
This is one of many exercises available from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises, a glossary, and chapters on supplies and laboratory techniques are also available at this site. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Echinodermata P, Eleutherozoa, Cryptosyringida, Echinozoa, Holothuroidea C, Dendrochirotacea sC, Dendrochirotida O, Sclerodactylidae F (Fig 9-26, 27-12, 28-62)
Echinodermata P
Echinoderms are secondarily radially symmetric deuterostomes whose ancestors were bilaterally symmetric. The adult radial symmetry is pentamerous with body parts occurring in fives or multiples thereof. Echinoderms have strong affinities with the ancestral trimeric deuterostomes especially in the tripartite organization of their coelomic cavities. Echinoderm larvae have the coelom divided into three regions, as is typical of the early coelomates, and these regions have important adult derivatives. All echinoderms are marine and benthic. About 6000 Recent species are known and the fossil record includes 13,000 extinct species.
An important echinoderm apomorphy is the water vascular system that in most taxa functions in support of locomotory tube feet but is also important in gas exchange, excretion, and feeding. The body wall includes a thick connective tissue dermis in which calcareous ossicles (little bones) are almost always an important component. These ossicles make up an endoskeleton that assumes different forms in different taxa. In most echinoderms calcareous spines of various sizes and shapes arise from the dermis and extend from the body surface and are alluded to by the name echinoderm (= spiny skin). The connective tissue is mutable and its consistency is under nervous control.
Excretion in echinoderms is accomplished by simple diffusion of metabolic wastes (ammonia) across thin permeable regions of the body wall. A variety of gas exchange structures, including the tube feet, is found in various echinoderms. A hemal system is present but its role in transport is still poorly understood and the chief transport system is the circulating fluid of the various coelomic compartments. The hemal system may be through transport system that delivers nutrients from the gut to these compartments for local distribution. The nervous system consists of two central intraepidermal nerve rings from which arise radial nerves to the periphery. Echinoderms are gonochoric and fertilization is usually external.
Eleutherozoa
Eleutherozoans are mobile echinoderms in which the oral surface is oriented against the substratum. A madreporite and locomotory tube feet are present. Polian vesicles and Tiedemann’s bodies may be present on the ring canal. Movable spines are present. Eleutherozoa includes all Recent echinoderms except for the sister taxon, Crinoidea.
Cryptosyringida
Cryptosyringida includes Ophiuroidea, Echinoidea, and Holothuroidea, all with closed ambulacra in which the radial nerve is internalized and protected by ossicles (Fig 28-20).
Echinozoa
Echinozoa, sister taxon to Ophiuroidea, consists of urchins and sea cucumbers. In these echinoderms the oral surface and ambulacra have expanded aborally until they enclose almost the entire body except for a small aboral periproctal region around the anus. A bony ring of ossicles surrounds the pharynx. The hemal system is better developed than in other echinoderms. The tube feet have ossicles
Holothuroidea C
The holothuroids, or sea cucumbers, are echinoderms in which the body is elongated along the aboral/oral axis. The body wall is thick and well developed. It consists of a nonciliated epidermis, connective tissue dermis, circular and longitudinal muscles, and a ciliated peritoneum. It is unusual in that its longitudinal muscles are in five longitudinal bands and the ossicles are microscopic plates scattered in the dermis. Abundant collagen fibers in the mutable dermis can, under nervous control, be cross-linked with each other to form a rigid skeleton that can scarcely be cut with a scalpel.
The gut is a simple tube extending from mouth to anus. Gas exchange is via diverticula of the posterior gut. Large branched tube feet encircle the mouth and are used for feeding. The hemal system is better developed than is typical in echinoderms and the coelom is large and spacious. The madreporite opens into the perivisceral coelom, rather than to the outside. Sea cucumbers are gonochoric and have a single gonad opening to the exterior via a gonopore at the oral end.
Dendrochirotida O
The body of dendrochirotes is more or less cucumber-shaped and the buccal tube feet are branched, or dendritic, and have no ampullae. Most taxa have a retractable introvert.
Laboratory Specimens
This exercise is written specifically for living specimens of the dendrochirote cucumber, Sclerodactyla briareus (= Thyone briaerius) but can also be used with other dendrochirotes. Sclerodactyla is a common, shallow water holothuroid of the east coast of North America from Massachusetts south to Florida and the Gulf of Mexico. Cucumaria frondosa is a northern dendrochirote occurring only as far south as southern New England and on the coasts of northern Europe (Fig 28-45). These two similar species belong to Dendrochirotacea and the dissection instructions can be used with either. Parenthetical comments point out differences between the two. If the anticipated dissection is to utilize preserved Cucumaria, it would be better to use the exercise written for that genus ( Invertebrate Anatomy OnLine ).
External Anatomy
Study a relaxed living cucumber in a dissecting pan immersed in 10% ethanol/seawater or isotonic magnesium chloride if living (or tapwater if preserved). The body is elongate with the mouth at the oral end and the anus at the opposite, aboral, end. A weak bilateral symmetry is evident against the underlying echinoderm radial symmetry. The aboral-oral axis is the long axis of the body and the axis of both bilateral and radial symmetry. The oral end is anterior and the aboral end is posterior.
The oral end bears the mouth and a circle of ten dendritically branched tentacles, or buccal podia (Fig 28-45). In both Sclerodactyla and Cucumaria the anterior end is an introvert that can be fully retracted into the body by a set of powerful retractor muscles. Threatened animals or those preserved without prior relaxation may have retracted introverts and the tentacles and mouth will not be apparent externally. If the tentacles are not visible in your specimen, try to evert them before you begin the dissection. Sometimes this can be accomplished by gently squeezing the body. If not, you will have to wait until you open the body cavity to see them.
Most cucumbers creep over the substratum or burrow into it. When creeping they keep the ventral surface in contact with the substratum and this surface is specialized for the purpose. The convex ventral surface has better developed tube feet, which are podia specialized for locomotion and usually have suckers. The dorsal surface of Sclerodactyla is concave and has poorly developed or absent suckerless podia.
An ambulacrum is a radial canal and its associated structures, including tube feet. Most echinoderms, holothuroids included, have five ambulacra radiating outward from the central ring canal (= water ring). Areas between the ambulacra are interambulacra.
The surface of the body bears small tube feet, or podia. The five ambulacra of many cucumbers, including Cucumaria, are marked externally by five longitudinal rows of these tube feet (Fig 28-45) but in Sclerodactyla they are scattered thickly over the body surface and the ambulacra, although present, are difficult or impossible to recognize from the outside. Three of the five ambulacra are associated with the ventral surface and are together known as the trivium , or sole (Fig 28-45). The two dorsal ambulacra are the bivium. Review the orientation of the cucumber and be sure you can find anterior (oral), posterior (aboral), dorsal, ventral, right, and left.
The longitudinal body wall muscles of cucumbers are concentrated in five strong longitudinal muscle bands which coincide with the ambulacra and can sometimes be seen externally. In species such as Sclerodactyla, in which the podia give no hint of the position of the ambulacra, the longitudinal muscles may.
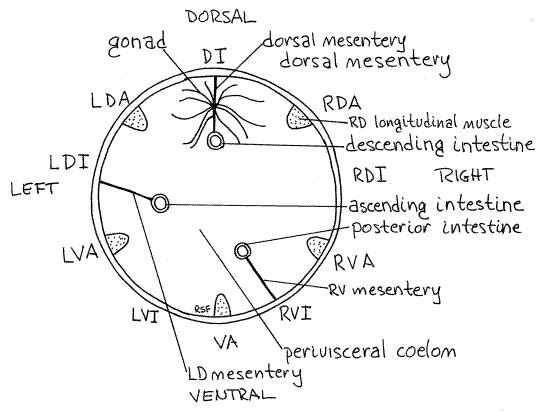
The dorsal midline coincides with an interambulacrum whereas the ventral midline is on an ambulacrum (Fig 1). If a cross section of a cucumber is compared with a clock face, the ambulacra are at roughly 2, 4, 6, 8, and 10 o'clock. The five ambulacra are referred to as right dorsal (2:00), left dorsal (10:00), right ventral (4:00), left ventral (8:00), and ventral (6:00). Similarly, the interambulacra are dorsal, right dorsal, left dorsal, right ventral, and left ventral (Fig 1).
Figure 1 A diagrammatic cross section of a typical cucumber showing the ambulacra, mesenteries, and gut loops. A = ambulacrum, I = interambulacrum, R = right, L = left, D = dorsal, V = ventral. Cuke11La.gif
The buccal podia are large, branched tube feet, one pair of which is associated with each ambulacrum. In Sclerodactyla the two tentacles of the ventral ambulacrum are much smaller than the others but in Cucumaria all are the same size.
Find the mouth at the center of the anterior end surrounded by a narrow peristomial membrane. A single small genital papilla , with the gonopore, lies on the dorsal midline between the trunks of the two dorsal buccal podia (Fig 28-45). The opposite, aboral end of the cucumber bears the anus at its center.
Internal Anatomy
" Perform the dissection with the specimen immersed in alcohol/seawater or isotonic magnesium chloride, if living, or tapwater if preserved. Use magnification as required. Open the body cavity by making a longitudinal incision from the anus anteriorly in the right dorsal interambulacrum (Fig 1). Use strong scissors for this and cut completely through the body wall without damaging the tissues within. Avoid cutting along or across the dorsal midline as a mesentery is located there. Extend the incision to the bases of the buccal podia. Pin the body wall aside so you can see the body cavity and its viscera. If your specimen is preserved, the body wall may be contracted, and you may have to make additional cuts before you can pin the wall against the wax. These cuts should be perpendicular to the original incision and should be made to the right of it.
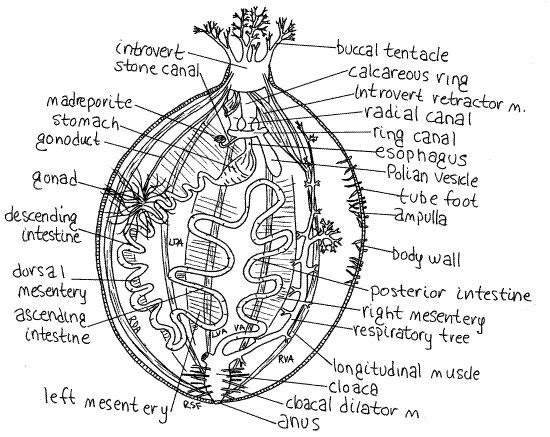
The body cavity, or perivisceral coelom, is large and spacious (Fig 2). Locate the major organs to use as landmarks. The white calcareous ring at the anterior end at the base of the tentacles (Fig 2) is a conspicuous feature. The yellow (Sclerodactyla) or orange (Cucumaria) gonad is a large tuft of long slender filaments that may fill much of the coelomic space, entangling and obscuring other structures. The gut is a translucent or opaque tube that loops its way from the anterior to the posterior end. Most of it is supported by mesenteries. The respiratory trees are large, delicate, translucent, branched tubes. The five pink, longitudinal muscle bundles can be seen on the inside of the body wall and mark the positions of the ambulacra. From the muscles arise five pink introvert retractor muscles that cross the body cavity to insert on the calcareous ring.
Body Wall
Look at the cut edge of the body wall and note the thick white layer of collagenous connective tissue. Most of the thickness of the wall is the connective tissue dermis. The thin epidermis lies outside the dermis.
Microscopic calcareous ossicles (Fig 28-47), which are present in the outer layer of the dermis of most cucumbers are rare in Sclerodactyla and Cucumaria.
>1a. Dermal ossicles can sometimes be found by digesting a bit of a buccal or an other podium in a drop or two of bleach on a slide. Make a wet mount and examine it with the compound microscope. <
The five longitudinal muscles are easily seen on the inner surface of the body wall. They are ambulacral in position and run from the posterior end of the body to insert on the middle of the calcareous ring. The introvert retractor muscles extend from them to the calcareous ring. Contraction of the retractor muscles pulls the calcareous ring into the coelom. The anterior end of the body, which is the introvert, is attached to the ring and consequently is pulled in also. Contraction of circular body wall muscles pressurizes the coelomic fluid and extends the introvert.
Calcareous Ring
The calcareous ring of 10 large ossicles, five ambulacral and five interambulacral, embedded in white connective tissue encircles the anterior end of the gut (Fig 2). It supports the ring canals of the water vascular and hemal systems, provides a mechanical base for the buccal podia and introvert, and is the site of insertion of the longitudinal body wall muscles and the introvert retractor muscles. The longitudinal muscles insert on the ambulacral ossicles with the introvert retractor muscles inserting posterior to the longitudinal muscles.
Figure 2. Dorsal dissection with the incision following the right dorsal interambulacrum. The left respiratory tree has been omitted and the right simplified for clarity. The hemal system is omitted. All introvert retractor muscles except the right ventral muscle are omitted. Abbreviations as in Fig 1. Based on Coe, 1912. cuke12La.gif
Coelom
The echinoderm coelom has many subdivisions of which the water vascular system and the trunk coelom are the two most conspicuous. The trunk coelom is subdivided into a large posteriorperivisceral coelom, which contains most of the viscera and is the major body cavity, and the peripharyngeal coelom around the calcareous ring. The calcareous ring and peripharyngeal coelom are enclosed in a mesenterial sac which, along with its contents, is the peripharyngeal bulb. The peripharyngeal coelom is not visible to you now but the perivisceral coelom and peripharyngeal bulb are.
The perivisceral coelom is incompletely partitioned by three longitudinal mesenteries. These are thin, transparent double sheets of peritoneum that extend from the body wall into the perivisceral coelom. They surround and support some of the visceral organs, especially the gut (Figs 1, 2). The dorsal mesentery runs along the dorsal midline on the dorsal interambulacrum (Fig 1). The left mesentery is in the left dorsal interambulacrum. The right mesentery is in the right ventral interambulacrum. Each of the three mesenteries is attached to a different region of the gut.
Coelomocytes of many types are present in the coelom. Those of the water vascular system often contain hemoglobin and are red in living animals.
Reproductive System
The single gonad is associated with the dorsal mesentery. Holothuroids are gonochoric but show no morphological sexual dimorphism and the tissues of the gonad must be examined microscopically to determine sex. In contrast to all other echinoderms, the reproductive system is bilaterally, not radially, symmetrical. The single gonad is a double tuft of long, spaghetti-like tubes whose free ends extend throughout much of the coelom. It is on the dorsal midline in the anterior perivisceral coelom (Fig 1, 2 28-50). The gonoduct (vas deferens or oviduct depending on sex) arises from the gonad and runs anteriorly along the dorsal midline. It is a distinct white tube extending to the genital papilla.
With the animal completely immersed in liquid, gently rake the filaments of the right tuft of the gonad together into a tidy bundle resembling a mop head. This will make it easier to observe its organization. Look at the anterior edge of the bundle where you will see much smaller, less heavily pigmented immature filaments. The number of filaments increases as the animal grows with new ones appearing anteriorly.
" The filaments of the gonad may be removed if they obscure the remaining viscera. Hold the clump of filaments together and use scissors to cut through the base of the tuft without damaging the dorsal mesentery, respiratory tree, or the associated gut. Discard the filaments. Don’t leave them in your pan or they will become entangled with everything else. Remove the left tuft using the same technique. If the mesentery makes it difficult to gather the second tuft into a bundle, you may have to remove it a few filaments at a time. Check to be sure the branches of the respiratory tree are not entangled in the filaments before you cut.
Digestive System
Move the two respiratory trees aside so you can see the gut tube, noting the close association between the left respiratory tree and the gut as you do.
The mouth opens into the pharynx which passes through the center of the calcareous ring where it cannot be seen. The esophagus exits the pharynx and calcareous ring to enter the perivisceral coelom (Fig 2). It soon joins the short, swollen, muscular stomach. (In Cucumaria the regions of the anterior gut are poorly defined and difficult to distinguish.)
The very long intestine exits the stomach and eventually empties into the cloaca at the posterior end of the coelom. The intestine makes two turns and is composed of three limbs. They are, in order, the descending anterior intestine, ascending anterior intestine, and the posterior intestine (= rectum). The descending anterior intestine runs posteriorly from the stomach and is attached to the body wall by the dorsal mesentery (Figs 1, 2, 28-50). In the posterior part of the body it reverses direction and becomes the ascending anterior intestine. This region of the intestine runs anteriorly attached to the body wall by the left mesentery. At the anterior end of the body cavity it reverses direction and becomes the posterior intestine. The posterior intestine runs posteriorly attached to the body wall by the right mesentery.
The posterior intestine joins the short, wide cloaca in the posterior end of the body (Fig 2). The cloaca is attached to the surrounding body wall by an abundance of threadlike radial cloacal dilator muscles. The opening of the intestine into the cloaca is small and kept tightly closed by the intestinal sphincter. The cloaca opens to the exterior via the anus which is closed by the anal sphincter. The respiratory trees are diverticula of the cloaca.
>1b. Open the cloaca with a longitudinal incision and find the opening of the posterior intestine and its sphincter in the anterior end. Also find the openings to the two respiratory trees on the lateral walls of the cloaca. <
Respiratory System
Gas exchange is accomplished chiefly by the two respiratory trees. These are hollow, highly branched diverticula of the cloacal wall. They extend into the perivisceral coelom and are bathed in coelomic fluid. The trees are anchored to the body wall by fine connective tissue threads and the left respiratory tree is intimately associated with the intestine.
>1c. Snip off a piece of the respiratory tree and place it in a small culture dish of seawater if alive (tapwater if preserved) and look at it with transmitted light on the stage of the dissecting microscope at high power (30X). Look for the thick layers of circular and longitudinal muscles in its walls. Find the cut end and verify that the branches are hollow. If your specimen is alive, the muscles will become active in the seawater and the tubules will writhe and bend as they contract. <
Holothuroids differ from other echinoderms in using muscles, rather than cilia, to generate the ventilating current. Expansion of the cloaca by contraction of the dilator muscles (with intestinal sphincter closed and anal sphincter relaxed) inspires water into the cloaca. With the intestinal and anal sphincters closed, circular cloacal muscles then contract to force the inhaled seawater into the respiratory trees. Gases are exchanged between coelomic fluid and seawater across the thin walls of the trees. Water is expelled by contraction of muscles in the walls of the trees (with intestinal sphincter closed, anal sphincter open).
Water Vascular System
The water vascular system of living Sclerodactyla is easily studied because its coelomocytes contain hemoglobin and the coelomic fluid is bright red. The color is lost in preserved material. The ring canal (= water ring) is located in the peripharyngeal bulb at the posterior end of the calcareous ring (Fig 2). It is a large, easily seen, membranous doughnut with thin, translucent walls. The esophagus passes through the hole in the doughnut. The small stone canal (= madreporic canal), arises from its anterior dorsal wall and extends a short distance into the perivisceral coelom to end at a small knotlike madreporite attached to the dorsal mesentery beside the gonoduct. Note that the madreporite opens into the perivisceral coelom rather than the sea. Its surface is covered by a ciliated epithelium through which pores open into the stone canal.
Also opening from the ring canal are the Polian vesicles, whose number varies with taxon. Sclerodactyla has 1-3 whereas Cucumaria has one. These thin-walled, hollow sacs are diverticula of the ring canal and are filled with water vascular system fluid. They apparently serve as reservoirs and are used to pressurize the system. (The Polian vesicle of Cucumaria arises on the left of the dorsal midline and is very long, extending to the posterior end of the coelom. It is highly distensible and may be slender when empty or grossly swollen when distended). Tiedemann's bodies are not present.
The five radial canals of the water vascular system arise from the anterior edge of the ring canal and extend anteriorly. They are large where they leave the ring canal and then widen into even larger translucent radial reservoirs between the ring canal and the posterior end of the calcareous ring. They then continue anteriorly, through the calcareous ring (out of sight) to emerge anterior to the calcareous ring. The canal then reverses direction and extends posteriorly on the body wall under a muscle bundle and supplies the tube feet via lateral canals. The radial canal is hidden by the muscle. The radial canals are supported by the ambulacral ossicles and the canals to the buccal podia lie between adjacent ambulacral and interambulacral ossicles.
The two ampullae of the two buccal podia of each radial canal flank the insertion of the retractor muscle of that particular ambulacrum and also flank the site of emergence of the radial canal from the peripharyngeal mass. The lateral canals to these two big ampullae exit the radial canal just after it (the radial canal) exits the radial reservoir.
>1d. Make a transverse section through the right dorsal ambulacrum and find the radial water canal between the longitudinal muscle and the body wall. <
In the body wall the radial canal gives rise to numerous lateral canals but these are hidden in the body wall musculature. Each lateral canal leads to the ampulla of a tube foot. The ampullae are small, elongate, transparent sacs on the inner surface of the body wall (Fig 2). In Sclerodactyla they are scattered over the entire surface, are not arranged in ambulacral rows, and look like little red worms (white in preserved specimens). Obviously, the lateral canals leading to the ampullae must be of varying lengths since the rube feet are varying distances from the canal. (In Cucumaria the ampullae are in neat rows on either side of the longitudinal muscle bundle of the ambulacrum and the lateral canals are all about the same length.)
The tube feet are cylindrical extensions of the body wall visible on the outside surface. Each contains a blind extension of the water vascular system connected to an ampulla and lateral canal.
Two buccal podia are associated with each of the five radial canals. Each has an ampulla with a lateral canal leading to it but the ampullae are quite large. They are long, translucent sacs anterior to the calcareous ring, near the insertions of the retractor muscles.
Hemal System
The holothuroid hemal system resembles that of other echinoderms but is better developed. No axial complex has been found in holothuroids. The dark, brownish hemal ring lies at the base of the ring canal (water ring) on its posterior face. It is a circular tube like the ring canal but it is smaller than the ring canal.
Five radial hemal canals arise from the hemal ring and accompany the radial water canals along the ambulacra. The origins of the radial hemal canals can be seen by opening the hemal ring and looking at its walls. They can also be seen in the cross section made earlier of the ambulacrum situated between the radial water canal and the integument.
The most conspicuous feature of the holothuroid hemal system is a pair of intestinal vessels and an elaborate set of vessels associated with them (Fig 28-51). The two intestinal vessels, the dorsal and ventral vessels, arise from the hemal ring and run posteriorly on opposite sides of the gut tube. A complex of capillaries connects the dorsal and ventral vessels across the gut. Larger transverse vessels connect the capillary beds of the descending anterior intestine with those of the ascending anterior intestine. This intricate network of fine vessels is a portal system known as the rete mirable (= miraculous net). It is better developed in aspidochirotacean cucumbers, such as Holothuria, than in Sclerodactyla and Cucumaria but its small transparent tubes are easily seen, even in these genera. The left respiratory tree is closely associated with the rete.
Nervous System
The peripheral nervous system arises from the nerve ring in the peristomial membrane. Five radial nerves exit the ring on the ambulacral axes. They accompany the radial water and radial hemal canals but, being intraepidermal, are peripheral to both. They can be seen in the cross section you made earlier to reveal the radial water canal. The dermis contains a nerve plexus. Chemosensory and mechanosensory receptors are present in the epidermis. Holothuroids exhibit a general sensitivity to light but no special photoreceptive organs are present.
References
Abbott DP. 1987. Observing Marine Invertebrates. Stanford Univ. Press, Stanford. 380.
Bullough WS . 1958. Practical Invertebrate Anatomy (2nd ed). MacMillan, London. 483p.
Coe WR. 1912. The echinoderms of Connecticut. Bull. Conn. State Geo. Nat. Hist. Surv. 19:1-152.
Edwards CL. 1910. Cucumaria frondosa. Zool. Jahrb. Abt. System 29:333-358, pl. 13.
Hyman LH . 1955. The Invertebrates: Echinodermata, vol. IV. McGraw-Hill, New York. 763pp.
Lawrence J. 1987. A Functional Biology of Echinoderms. Johns Hopkins, Baltimore. 340p.
Reid WM. 1950. Thyone briaerius, pp 541-546 in Brown FA. (ed.) Selected Invertebrate Types. Wiley, New York. 597p.
Ruppert EE, Fox RS, Barnes RB. 2004. Invertebrate Zoology, A functional evolutionary approach, 7 th ed. Brooks Cole Thomson, Belmont CA. 963 pp.
Smiley S. 1994. Holothuroidea, pp. 401-471 in Harrison FW, Chia FS (eds.). Microscopic Anatomy of Invertebrates vol. 14 Echinodermata. Wiley-Liss, New York. 510p.
Supplies
Dissecting microscope
Compound microscope
Slides, coverslips
Dissecting pan
Living or preserved cucumber
Isotonic magnesium chloride or 7% ethanol (non-denatured) in seawater for living cukes
# 4 stainless steel insect pins