Invertebrate Anatomy OnLine
Romalea microptera ©
Eastern Lubber Grasshopper
6oct2006
Copyright 2001, 2004 by
Richard Fox
Lander University
Preface
This is one of many exercises available from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises can be accessed by clicking on the links to the left. A glossary and chapters on supplies and laboratory techniques are also available. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Arthropoda P, Mandibulata, Tracheata, Hexapoda SC, Insecta C, Dicondylia, Pterygota, Metapterygota, Neoptera, Orthopterodea SO, Orthoptera O, Caelifera sO, Acrididea iO, Acridoidea SF, Romaleidae F, Romaleini T, (Fig 16-15, 20-14, 20-15, 21-23)
Arthropoda
Arthropoda, by far the largest and most diverse animal taxon, includes chelicerates, insects, myriapods, and crustaceans as well as many extinct taxa. The body is segmented and primitively bears a pair of jointed appendages on each segment. The epidermis secretes a complex cuticular exoskeleton which must be molted to permit increase in size. Extant arthropods exhibit regional specialization in the structure and function of segments and appendages. The body is typically divided into a head and trunk, of which the trunk is often itself divided into thorax and abdomen.
The gut consists of foregut, midgut, and hindgut and extends the length of the body from anterior mouth to posterior anus. Foregut and hindgut are epidermal invaginations, being derived from the embryonic stomodeum and proctodeum respectively, and are lined by cuticle, as are all epidermal surfaces. The midgut is endodermal and is responsible for most enzyme secretion, hydrolysis, and absorption.
The coelom is reduced to small spaces associated with the gonads and kidney. The functional body cavity is a spacious hemocoel divided by a horizontal diaphragm into a dorsal pericardial sinus and a much larger perivisceral sinus. Sometimes there is a small ventral perineural sinus surrounding the ventral nerve cord.
The hemal system includes a dorsal, contractile, tubular, ostiate heart that pumps blood to and from the hemocoel. Excretory organs vary with taxon and include Malpighian tubules, saccate nephridia, and nephrocytes. Respiratory organs also vary with taxon and include many types of gills, book lungs, and tracheae.
The nervous system consists of a dorsal, anterior brain of two or three pairs of ganglia, circumenteric connectives, and a paired ventral nerve cord with segmental ganglia and segmental peripheral nerves. Various degrees of condensation and cephalization are found in different taxa.
Development is derived with centrolecithal eggs and superficial cleavage. There is frequently a larva although development is direct in many. Juveniles pass through a series of instars separated by molts until reaching the adult size and reproductive condition. At this time molting and growth may cease or continue, depending on taxon.
Mandibulata
Mandibulata includes arthropods in which the third head segment bears a pair of mandibles. As currently conceived this taxon includes myriapods, hexapods, and crustaceans. Appendages may be uni- or biramous and habitats include marine, freshwater, terrestrial, and aerial.
Tracheata
Myriapods and hexapods share tracheae and a single pair of antennae and are sister taxa in Tracheata. Crustaceans, which have gills and lack tracheae, are excluded and form the sister group.
Hexapoda
The body is divided into three tagmata; head, thorax, and abdomen. Appendages are uniramous and a single pair of antennae is present. Three pairs of legs and two pairs of wings are found on the thorax of most adults. Hexapod legs are uniramous although there is increasing evidence that they evolved from multiramous appendages of their ancestors. Gas exchange is accomplished by trachea. Excretory organs are Malpighian tubules and the end product of nitrogen metabolism is uric acid. There is relatively little cephalization of the nervous system. Insects are gonochoric with copulation and internal fertilization.
Insecta
Most hexapods are insects. A few hexapod taxa (orders) lack wings and have primitive mouthparts recessed into the head and belong to Entognatha, the sister taxon of Insecta. Insects have ectognath mouthparts and the adults (imagoes) of most taxa have wings.
Pterygota
Pterygotes are the winged insects. These insects are derived from a winged common ancestor. Adults of most taxa have wings although they have been lost in some.
Orthoptera
Orthoptera includes 20,000 species of grasshoppers, crickets, locusts, katydids, and their relatives. These are mostly large insects with an enlarged pronotum. The hind femora are large and adapted for jumping. Females have a large ovipositor but male genitalia are not visible externally. Most are herbivores.
Natural History
Lubbers are tropical short-horned grasshoppers of the family Romaleidae. The family is largely restricted to Central and South America but one species, Romalea microptera (= R. guttata), occurs in the southeastern United States where its range once extended from Florida to Texas and north to Tennessee and the Carolinas. Several other lubber species are confined to arid regions of the western United States but R. microptera is adapted to moister habitats and is the only species in the east.
Until recently, isolated populations of R. microptera were present in Tennessee, Arkansas, South Carolina, Florida, and Louisiana. Eastern lubbers remain abundant in Florida, especially in the Everglades, but at present the only confirmed population remaining outside of Florida is in South Carolina. Anyone having knowledge of extant R. microptera populations, other than those in Florida, may communicate that information to Dr. Doug Whitman of Illinois State University at dwwhitm@ilstu.edu . Habitat destruction and pesticides are probably responsible for the decline of this species. Although its distribution is patchy, where present the eastern lubber can be very common in the summer.
The lubber family (Romaleidae) is characterized by the presence of a pair of strong spines at the tip of the hind tibia. One spine is medial, the other lateral. Lubbers (and some other grasshoppers) also have a prominent midventral tooth-like process on the first thoracic segment, between the first pair of legs. The common name “lubber” alludes to the stocky, heavy-bodied appearance of members of this family. The Merriam-Webster Collegiate Dictionary defines lubber as a “big clumsy fellow”.
The wings of most lubber grasshoppers are rudimentary and incapable of flight. Romalea microptera has reduced, non-functional wings and is flightless (microptera = small wing). Consequently its dispersive ability is weak and there is little gene flow between populations. Grasshoppers have chewing mouthparts and lubbers use theirs to feed on almost any green vegetation but they prefer lilies (in the family Lilaceae). Members of this plant family synthesize allelochemicals to protect themselves from herbivores but these compounds are ineffective against lubbers. Instead, lubber grasshoppers save toxins from the lilies they ingest and use them in their own defense.
Like many chemically protected organisms lubbers are aposematically colored to discourage predation. Nymphs are usually jet black with bright orange, yellow, or red markings, including a distinct middorsal red stripe. (A few individuals may be pale pinkish orange with darker red markings.) In contrast, adults are cryptic with black, yellow, and brown markings. The hind wings, however, are hot pink with a black border but they are normally hidden by the forewings, which have a faint pink blush. (An uncommon black color morph is found on the Gulf of Mexico coast. Adults of this morph are black with yellow markings.)
When threatened, an adult lifts the forewings and spreads the hindwings to display its startling pink and black pattern which may resemble an eye to a would-be predator. It also hisses and releases a repugnatorial substance from glands in the thorax. Like other grasshoppers it releases a repellant oral secretion (known to children as “tobacco juice” that also contains toxins from its diet. It has other toxins in the blood. A single bad experience with a protected lubber is sufficient to cause a predator to avoid all lubbers in the future.
Romalea is gonochoric but with little sexual dimorphism other than a marked disparity in size. Females (about 75 mm) are larger than males (about 55 mm) and have a short ovipositor which is lacking in males.
Like other orthopterans lubbers are paurometabolous (Fig 21-13). Juveniles resemble adults and undergo a gradual increase in size, but not shape, from juvenile to imago. Lubbers have an annual life cycle and females inject eggs into the ground in late summer. During mating and until oviposition the smaller male rides the female to guard against insemination by rival males. Adults die after oviposition. Eggs enter diapause, overwintering in the ground to hatch in the spring. First instar nymphs appear in April in South Carolina and are black and red as described above. In Florida nymphs may appear as early as February. Lubbers have four nymphal instars, all with similar pigmentation and appearance except that they become progressively larger as spring and summer progress. The final molt, in summer, transforms the final juvenile instar into the winged and sexually mature imago (adult). In 2006, in Greenwood, South Carolina, the first imagoes were recorded on 24jun. None was seen after 10sep.
Early instar nymphs remain together in sibling groups consisting of the individuals from a single oviposition. Later instars and imagoes, however, leave these aggregations and go their separate ways.
Natural history information is from Whitman (2006), Eisner (2003), and from observations of the Greenwood, South Carolina population.
Laboratory Specimens
Because they are relatively primitive, more or less typical insects, grasshoppers are often used in introductory biology laboratories as an introduction to insect or arthropod anatomy. In North America, the eastern lubber grasshopper, Romalea microptera (= R. guttata), is the species usually employed for this purpose because it is large and readily available. Adults are 50-75 mm in length and are easily dissected and provide a good example of general, relatively unspecialized insect anatomy. This is usually the species provided by biological supply companies although other grasshoppers are sometimes found in their shipments. (Any specimens with large wings are not Romalea.)
Romalea, like most animals, is best studied using anesthetized living, or freshly sacrificed specimens. Preserved animals are less satisfactory. Unfortunately, the lubber life cycle is such that living adults are not usually available during the academic year when they are needed by most invertebrate zoology courses and consequently it is almost always studied using commercially provided preserved material. The cricket, Acheta domestica, in contrast, is available alive and inexpensively at bait shops throughout the year and is recommended as a substitute. Dissection instructions for Acheta are available at Invertebrate Anatomy OnLine.
Conduct the study using a dissecting microscope. Living or preserved specimens can be used. This description is based on fresh, unpreserved specimens. References to color usually are not applicable to preserved material. Anesthetization of living specimens is optional for the study of external anatomy. Anesthetization can be accomplished with carbon dioxide or chloroform. For the study of internal anatomy, living specimens can be anesthetized as above or killed by exposure to fumes from a cotton ball dampened with ethyl acetate in a covered culture dish.
External Anatomy
Tagmata
Examine a male or female Romalea under magnification. The arthropod body is composed of a linear series of segments, some or all of which bear paired jointed segmental appendages. Most arthropods, including insects, are heteronomous, with their segments and their appendages variously modified and grouped in regions, or tagmata, specialized for different functions. The alternative homonomous condition, with all segments and appendages alike over the length of the body, is primitive but, among Recent taxa, is approached only by a few crustaceans.
Figure 1. Lateral view of an adult male lubber grasshopper from Greenwood, South Carolina. Orthop19La.gif
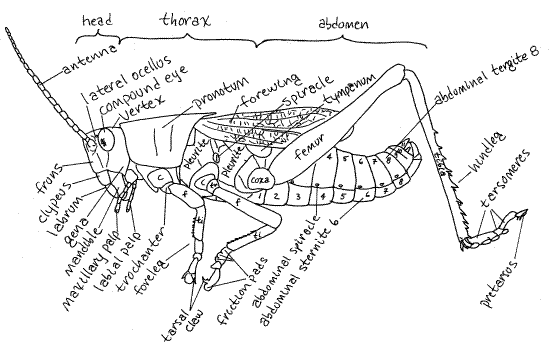
The three tagmata of the insect body are the anterior head, middle thorax, and posterior abdomen (Fig 1, 21-1C). The head, which shows few external signs of segmentation, bears the eyes, antennae, mouth, and mouthparts. Its major functions are sensory reception and feeding. The thorax, whose primary function is locomotion, is larger and bears three pairs of legs and, typically, two pairs of wings. The abdomen is the largest tagma and is conspicuously segmented. It houses most of the digestive, excretory, and reproductive viscera and its appendages, when present, are specialized for copulation or oviposition. Most abdominal segments lack appendages and those present are highly modified. The external genitalia, at the posterior end of the abdomen, are highly modified segmental appendages.
Head
The head is enclosed in a hard, heavily sclerotized, unsegmented, exoskeletal head capsule, or epicranium. The mouthparts are attached ventrally to the capsule, a condition known as hypognathus. The mouth, which you will see later, is located on the ventral surface of the head and is surrounded by the mouthparts. These mouthparts enclose a space, the preoral cavity (Fig 21-7), from which the mouth opens. The head is more or less ovoid in shape with a nearly vertical anterior surface.
Figure 2. En face view of Romalea. Orthop20L.gif
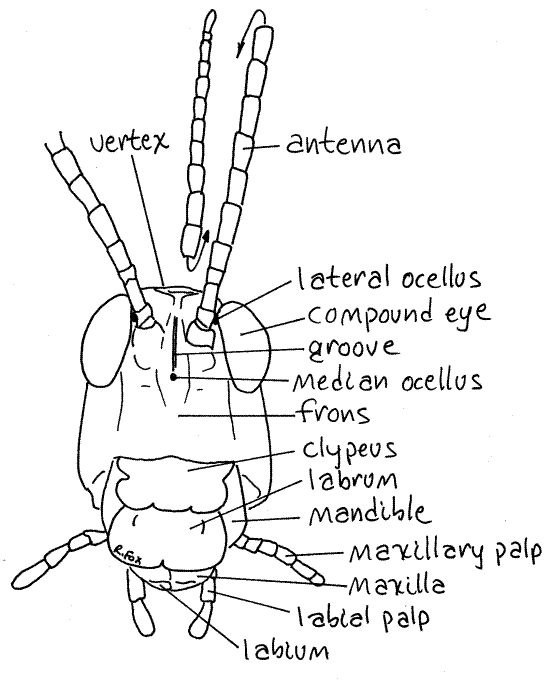
Note the composition of the head capsule. Most of the head is enclosed in a single hard piece of exoskeleton known as the epicranium. This is composed of the fused sclerites of the head segments. It covers the dorsal anterior and lateral aspects of the head and is divided into regions. The region of the epicranium anterior to the compound eyes is the frons (Fig 1, 2). The side, or cheek, posterior to the compound eye is the gena (Fig 1). Ventral to the frons is the clypeus (Fig 2, 4). The unpaired clypeus (CLIP ee us) articulates with the epicranium. The short lateral edges of the clypeus are notched. The vertex is the top of the capsule, dorsal to the frons and between the compound eyes. The vertex extends anteriorly above the bases of the antennae as a prominent, pointed, concave rostrum (Fig 3). The posterior wall of the head capsule is penetrated by a large aperture, the foramen. Through it pass the organ systems, such as nerve cord, esophagus, salivary ducts, and musculature, connecting the head with the thorax. In an intact specimen the foramen is filled with soft tissue but you can see its circular outline. The occiput (occipital arch) is a narrow band forming the posterior edge of the head capsule arching dorsally over the foramen.
Figure 3. Dorsal view of the head and prothorax of a female Romalea from Greenwood, SC. Orthop36L.gif
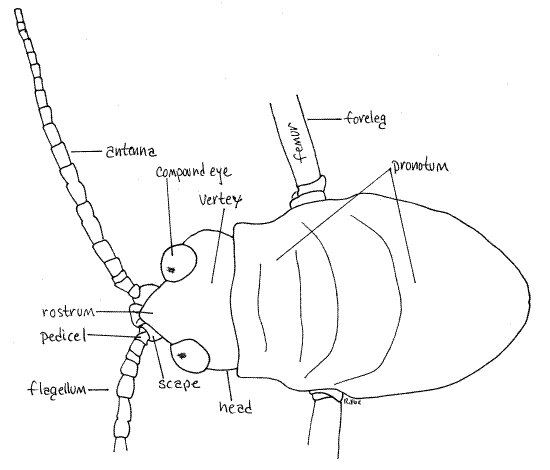
A pair of large multiarticulate, sensory antennae is attached in deep sockets on the antero-dorsal corners of the head (Fig 1, 2, 3, 21-1A,B). The antenna consists of two basal articles, the proximal scape and distal pedicel, to which is attached the long multiarticulate filamentous flagellum (Fig 3). The antennae are the first pair of segmental appendages of the insect head and are innervated by the deutocerebrum of the brain. Tracheates have only one pair of antennae in contrast with crustaceans which have two and chelicerates which have none.
A large, bulging compound eye is located dorsolaterally on each side of the head just posterior to the base of the antenna (Fig 1, 2, 21-1). Examine the surface of a compound eye and note that it is composed of an uncountable number of small light-receiving units known as ommatidia. The surface of the eye is a specialized, transparent part of the exoskeleton divided into tiny hexagonal cuticular lenses, or corneas, one for each ommatidium.
In addition to the two compound eyes there are three tiny, simple eyes, or ocelli (Fig 2, 21-1A). These are not composed of ommatidia. Two of them are lateral ocelli located between the compound eyes and the antennae (Fig 2). The unpaired median ocellus is located on the anterior midline of the head in a pit on the vertical ridge between the two antennae (Fig 2).
Figure 4. En face view of the clypeus and labrum of Romalea. Orthop21L.gif
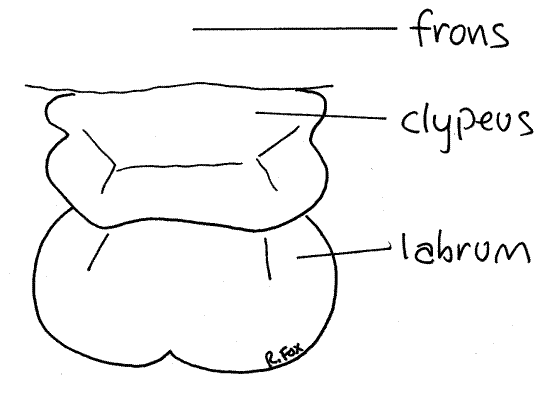
The labrum, or upper lip, is also located on the vertical anterior face of the head capsule (Fig 1, 2, 4, 21-1). It is a large movable plate, equipped with muscles, ventral to the clypeus. It bears a small median notch on its ventral border and a median groove on its anterior surface. Some entomologists believe the labrum to be derived from a fused pair of anterior appendages and that the notch and groove represent the line of fusion between the fused right and left appendages.
The labrum is the anterior wall of the preoral cavity and it covers the more posterior mouthparts. Its hidden posterior surface bears abundant setae, or bristles. With fine forceps lift the labrum to demonstrate its mobility and to reveal the preoral cavity and the remaining head appendages. The labrum is innervated by the tritocerebrum indicating it is the homolog of the crustacean second antenna and supporting the hypothesis that it is a segmental appendage.
Figure 5. Anterior (left) and posterior (right) views of the left mandible of Romalea. Orthop22L.gif
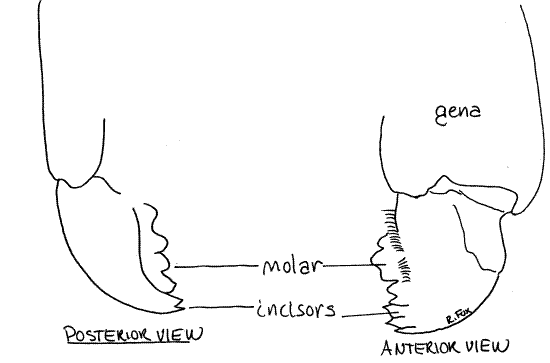
Immediately posterior to the labrum is a pair of mandibles (Figs 1, 2, 5, 21-1). In grasshoppers these massive mouthparts are adapted for biting and chewing. Their heavily sclerotized and strongly toothed median surfaces are apparent when the labrum is moved aside. The mandibles are segmental appendages.
The toothed, median, cutting surface of the mandible includes a distal (ventral) incisor of sharp shearing teeth and a proximal molar of heavier grinding teeth (Fig 5). You can see these by lifting the labrum. You may later want to remove the mouthparts on one side for closer study but do not do so now.
The mandible articulates with the ventral edge of the posterior epicranium (gena). The mandibles lie on either side of the mouth and are the sides of the preoral cavity from which the mouth opens dorsally. You will not see the mouth at present.
If your specimen is alive and unanesthetized, it may move its mandibles as you watch. They are operated by powerful muscles with the motion entirely in the transverse plane. Grasshopper mandibles are dicondylic, meaning they are hinge joints, articulating with the epicranium at two points, known as condyles, thus limiting the range of motion to a single plane. A monocondylic articulation, on the other hand, would be a ball and socket joint with a much greater range of motion. Grasp a mandible with fine forceps and move it to demonstrate its motion.
The soft, fleshy hypopharynx extends into the preoral cavity as a fold of body wall ventrally from the head capsule just posterior to the mouth. This unpaired, tongue-like structure is not a segmental appendage. It divides the preoral cavity into an anterior cibarium, from which the mouth opens, and a posterior salivarium, into which the duct of the salivary glands open (Fig 21-7).
Figure 6. The left maxilla of Romalea. Orthop23L.gif
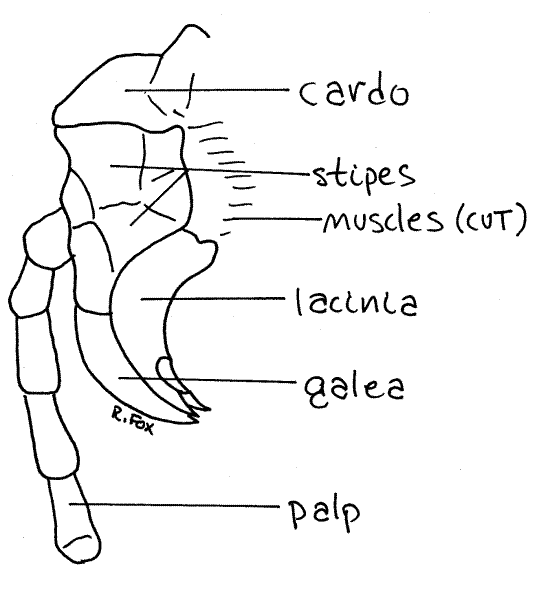
The appendages immediately posterior to the mandible are the two maxillae (Figs 1, 6, 21-1). These also articulate with the gena of the epicranium. Each maxilla consists of two basal articles. One, the cardo, articulates with the gena. The other, the stipes, articulates with the cardo. From the stipes arises a spoon-shaped lateral galea and a toothed medial lacinia.
The galea and lacinia curve medially, just posterior to the mandibular incisor. The galea hides the lacinia from lateral view. You can roll the maxilla aside on its articulation with the gena to reveal these parts. A large, conspicuous, filamentary, and multiarticulate maxillary palp also arises from the stipes but on its lateral side (Figs 1, 6). The palp is sensory.
The last pair of head appendages are fused to form the labium, or lower lip (Figs 1, 7, 21-1). The labium is easily seen in its entirety by looking at the posterior surface of the ventral head. It forms the posterior boundary of the preoral cavity. The labium articulates with a slender U-shaped arch of exoskeleton (a modified sternite) known as the gula (Fig 7).
Figure 7. The labium of Romalea. Orthop24L.gif
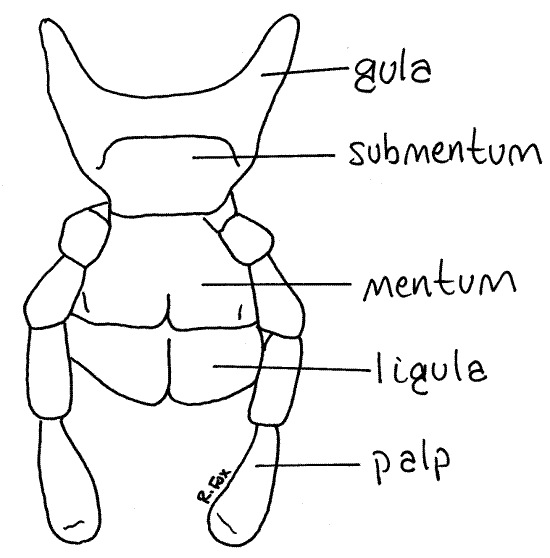
The arms of the gula extend anteriorly to the posterior edge of the gena. The basal portion of the labium consists of a small, more or less, rectangular, proximal submentum, which articulates with the gula, and the larger, more distal mentum. The anterior edge of the mentum is cleft medially, reminding us that the labium is formed evolutionarily of two fused segmental appendages. The mentum bears two large cupped ligulae. The lateral borders of the mentum each bear a filamentous, multiarticulate, sensory labial palp.
The labium forms the posterior wall of the preoral cavity. Push the labium posteriorly and look into the preoral cavity. Its anterior wall is the labrum. The mandibles and maxillae are the sides. Inside the cavity you will see the large hypopharynx extending from the ventral wall of the head immediately posterior to the mouth. It is an unpaired fold of the body wall and is not a segmental appendage. The mouth lies between the base of the hypopharynx and the labrum.
Thorax
The heavily sclerotized head and thorax are connected by a short narrow neck (Fig 1), or cervix, whose integument is lightly sclerotized and consequently soft and flexible. The neck and its leathery integument are easiest to see ventrally but are hidden dorsally by the head and thorax. A few hard, sclerotized plates, the cervical sclerites, are embedded in its integument.
The insect thorax consists of three segments, each of which bears a pair of legs. The legs are segmental appendages. Two of the thoracic segments typically bear a pair of wings, which are complex folds of the body wall and are not segmental appendages. Wings, present only in imagoes, are present in most insect orders.
The three thoracic segments are, in order from anterior to posterior, the prothorax, mesothorax, and metathorax (Fig 1). The meso- and metathoraces are fused rigidly to form an inflexible box housing the flight muscles and bearing the two pairs of wings. Together they are referred to as the pterothorax in reference to the wings they bear (pter = wing). The prothorax is joined to the pterothorax and to the head by flexible articulations.
A typical arthropod segment is enclosed in four exoskeletal plates, or sclerites, that form a ring around the segment (Fig 16-1B). These are the dorsal tergite, ventral sternite, and two lateral pleurites. These sclerites are typically joined together by flexible articulating membranes but sometimes they are fused rigidly together. This primitive pattern often undergoes some modification in modern arthropods, including grasshoppers.
Prothorax
The grasshopper prothorax is covered by an unusually large shieldlike tergal plate called the pronotum (Fig 1, 3, 21-1C). (The tergites of the thorax are called nota, rather than tergites. Fortunately, thoracic sternites and pleurites do not have special names.) The pronotum covers the prothorax and partly overhangs the mesothorax. The prosternite, located ventrally, is much smaller and bears a conspicuous median prosternal spine, by which may be recognized.
The forelegs (Fig 1, 21-1C), or prothoracic legs, articulate with the prothorax. The pleurites of the prothorax, known as propleurites, are small and located between the prosternite and the pronotum. They are fused with both the prosternite and the pronotum. Together the prosternite and propleurites form sockets for the articulation of the forelegs. The propleurite forms the dorsal edge of the socket whereas the prosternite forms the ventral edge.
Note the flexible, unsclerotized cuticle forming an articular membrane between the base of the leg and the socket. Rotate the leg around the articulation to demonstrate its mobility.
Primitively, each body segment bears a pair of lateral openings, or spiracles, for the intake of air into the tracheal respiratory system. One such spiracle is found on each side of the prothorax. It is located laterally near the border between the prothorax and mesothorax in the soft unsclerotized cuticle under the posterior edge of the pronotum. The prothoracic spiracles are difficult to see because they are hidden by the pronotum. The more posterior spiracles are easier to locate.
Pterothorax
The pterothorax comprises the mesothorax and metathorax. Each segment bears a pair of wings and a pair of legs. The wings of the mesothorax are the forewings, or wing covers (Fig 1), and those of the metathorax are the hindwings. The mesothorax bears the middle legs (Fig 1, 21-1C), or mesothoracic legs. The metathorax bears the hindlegs, or metathoracic legs.
Dorsally the two segments of the pterothorax are covered by the two sclerotized tergites known as the mesonotum and metanotum. Similarly the ventral surfaces are protected by the mesosternite and metasternite. Note the manner in which sternites join together along their lines of fusion. The metasternite is dovetailed into the mesosternite. The sternite of the first abdominal segment resembles the thoracic sternites and is dovetailed into the posterior edge of the metasternite.
Deep invaginations at the corners of the dovetails are apophyseal pits marking the position of important sternal apophyses, or apodemes, extending from the exoskeleton into the interior of the pterothorax to function as an endoskeleton.
The sides of the pterothorax are composed of sclerites derived from the pleura of ancient insects. The pleurite on each side of both pterothoracic segments is divided into two smaller plates. The legs articulate between the ventral edge of the pleurites and the sternites. The wings arise between the dorsal edge of the pleurites and the nota (tergites).
A spiracle is situated laterally between the pleura of the metathorax and mesothorax (Fig 1). Note that its opening is guarded by two movable lips, or valves. The opening is a straight slit between the two lips. The spiracle opens into the tracheal system.
Legs
Study one of the middle legs. This appendage, which is typical of insect legs, is composed of a linear series of units, or articles. Each article consists of a sclerotized exoskeletal cylinder which contains muscles, tendons, blood, connective tissue, and nerves. Note that while most of the limb exoskeleton is rigid and heavily sclerotized, some parts are flexible and unsclerotized. The unsclerotized regions are the movable articulations between successive articles (Fig 16-1D). Look at some of the articulations under magnification and find these flexible articular membranes.
The first (proximal) article is the leg is the coxa (Fig 1, 21-1E). It is short, wide at the base, narrow distally, and articulates by a broad articulating membrane with the socket formed by the pleurite and sternite of the mesothorax.
Articulated with the distal end of the coxa is the even shorter, cylindrical trochanter. The trochanter is fused immovably with the long femur. The femur is one of the two long articles of the leg. The tibia, which is the second of the long articles, articulates with the distal end of the femur.
Pay particular attention to the hard parts of the articulation between the femur and tibia. They form a dicondylic hinge joint similar to that in your elbow and knee (but exoskeletal rather than endoskeletal, of course). Such a joint restricts motion to a single plane. You encountered another dicondylic joint between the mandible and epicranium. The tibia is long and slender with teeth on its margins.
Next in line along the length of the leg is the tarsus composed of two subdivisions, or tarsomeres (Fig 1, 21-1E). The number of tarsomeres varies among insects. The proximal tarsomere bears three soft, unsclerotized friction pads on its surface to prevent slipping of the tarsus on the substratum. The distal tarsomere is much shorter and has a single friction pad.
The final article of the leg is the pretarsus. It extends from the distal end of the tarsus and bears a pair of sclerotized tarsal claws and a single soft friction pad.
The forelegs are nearly identical to the midlegs but the hindlegs are much large. and are responsible for the characteristic jumping (saltational) locomotion of grasshoppers. The large coxa of the hind leg is fused with the trochanter and the femur and tibia are much elongated. In addition, the femur is expanded proximally to accommodate the major jumping muscles. This expansion gives it a club or drumstick shape.
Wings
The two pairs of wings are evaginations or folds of the cuticle and body wall of the two segments of the pterothorax. Each wing is a double layer of body wall consisting largely of cuticle but underlain and secreted by epidermis. The characteristic "veins" of the wings are thickened tubes of exoskeleton involved in support. Their primary function is structural but they contain epidermis (which secretes the cuticle), nerves, and blood.
The wings of the flightless Romalea microptera are small and are not used for flight. The forewings of all grasshoppers, including Romalea, are heavy and form a pair of wing covers to protect the more delicate hindwings. Those grasshoppers capable of flight use the hindwings, and not the forewings, for flying.
Lift and spread the wing covers (forewings) and examine them with magnification. Note the heavy longitudinal veins and the abundance of small, branching and anastomosing veins arising from them. Make a similar examination of the hindwings.
If you are studying an immature grasshopper, the wings will be absent or represented by small thick wing pads whose size depends on the maturity of the juvenile. Wing pads increase in size with each molt and become wings with the final molt. Wings are present only in adult insects.
Abdomen
The grasshopper abdomen consists of 11 segments and extends posteriorly from the thorax (Fig 1, 21-1F). It is the largest of the tagmata and most of its segments have no appendages. Its segmentation is obvious externally, moreso than that of the other tagmata, and more like that of the ancestral arthropods.
With the exception of those at the posterior end, grasshopper abdominal segments are easily recognized and counted (Fig 1, 21-1F). Numbered from anterior to posterior, abdominal segments 1-7 are similar to each other and are typical, unspecialized insect segments. These are the pregenital segments and none bears appendages. Segments 8-9 are reduced and modified and may bear appendages. These are the genital segments and may have appendages modified as external genitalia. The gonopore is on segment 8 but it is hidden by the genitalia. Segments 10-11 are the postgenital segments.
Typically each segment is covered by a large dorsal tergite and a ventral sternite (Fig 1, 16-1B). The two articulate laterally via a flexible, narrow articular membrane. Abdominal sclerites are not as heavily sclerotized as are those of the thorax and head. Eleven tergites are present but some of the posterior segments lack sternites and, as a consequence, there are only eight sternites in females and nine in males (Figs 7, 8). The pleurites are reduced and fused to the ventral edges of the tergites.
Most abdominal segments are similar to each other but the anterior and posterior ones show some specializations. The first abdominal segment is fused rigidly with the thorax. You have already seen its sternite dovetailed into the metasternite. Its tergite is mostly hidden by the base of the hindwings.
Each side of tergite 1 bears a large aperture covered by a thin cuticular tympanic membrane (= tympanum or eardrum). These are auditory organs for sound transduction. Some species of lubber grasshoppers have stridulating organs with which they produce sound.
The spiracle of the first abdominal segment is located on the anterior edge of the tympanum. This spiracle opens into a large air sac attached to the inner surface of the tympanum.
Segments 2-7 are similar. The fifth abdominal segment is typical of the pregenital segments (Fig 1, 21-1F). Its tergite forms a large arch over the dorsum and its sternite forms a broad band across the belly. Both sclerites are joined to each other and with adjacent sternites and tergites by flexible articular membranes. A spiracle is located at the extreme anterior ventral corner of the tergite.
The posterior abdominal segments are associated with the external genitalia and differ from other segments and between males and females. Some bear modified segmental appendages. The large anus is on segment 11 ventral to tergite 11 (tergite 11 is the epiproct). It is covered dorsally by the single epiproct and flanked by two paraprocts. The ovipositor of females and penis (= aedeagus) of males are ventral to the anus. The posterior abdomens of males and females differ and must be studied independently. Exchange specimens with another student in order to study the opposite sex.
Female
Externally, female segment 8 is similar to the preceding segments except that its sternite is known as the genital plate (Fig 8, 9). The egg guide is a triangular, median process of the genital plate. Segments posterior to 8 are reduced and lack sternites (Fig 21-11B). The tergites of segments 9 and 10 are reduced and fused laterally although they remain separate dorsally (Fig 8). The 11 th tergite, or epiproct, is rather large and it forms a triangular dorsal shield rather than an arch over the body. Its pointed end projects posteriorly and covers the anus. Associated with it is a lateral paraproct on each side, ventral to the epiproct. The paraprocts are probably the sternites of segment 11.
Between the 11 th tergite and each of the paraprocts is a small cercus. The cerci are the segmental appendages of segment 11. In some orthopterans, such as crickets, they are long antenniform sensory organs but in Romalea they are very short.
The female external genitalia are modified segmental appendages belonging to segments 8 and 9. Together they form the short robust ovipositor extending posteriorly from the abdomen.
Figure 8. Lateral view of female abdomen. Orthop25L.gif
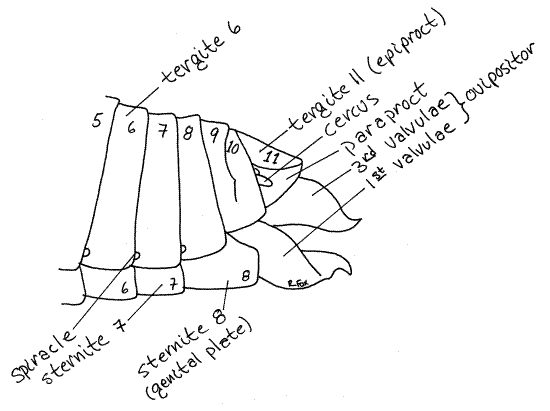
Figure 9. Posterior view of the external genitalia of a female Romalea. The first and third valvulae are reflected ventrally and dorsally, respectively. Orthop37L.gif
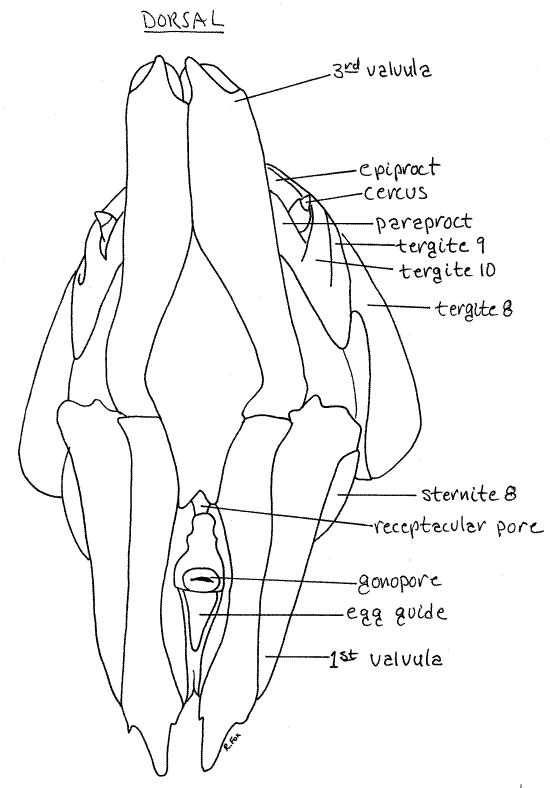
The ovipositor consists of three pairs of processes, called valvulae, which are used by the female to insert eggs into the ground. Ventralmost is the pair of large, heavily sclerotized first valvulae (= ventral valves)(Fig 8, 9). Dorsally is a similar pair of sclerotized third valvulae (= dorsal valves). The first and third valvulae are conspicuous externally. The inconspicuous, membranous second valvulae (= inner valves) are hidden from view by the first and third and will not be seen. The gonopore lies between the first valvulae (Fig 9). The space enclosed by the epiproct, paraprocts, genital plate and ovipositor is the genital chamber.
The right and left first and third valvulae together form a hollow shaft, or ovipositor, consisting of four parts, that is used to penetrate the soil. The eggs pass from the gonopore, through the lumen of the shaft, assisted by the second valvulae, and are deposited in masses in the soil.
The first valvulae are derived from the appendages of segment 8 whereas the second and third are the appendages of segment 9.
Male
The male posterior abdomen is simpler than that of the female (Fig 10, 11, 21-12B). Segment 8 resembles segments 2-7 and has an unmodified sternite. Tergites 9-10 are fused ventrally and separate dorsally as in females. Segment 9 possesses a large sternite, the subgenital plate that extends posteriorly ventral to segments 10-11. It forms a cup enclosing the male external genitalia. Protected and hidden by the plate is the soft (largely unsclerotized) eversible penis (= aedeagus). Although referred to as “external” genitalia, the penis is not visible externally without moving the paraprocts and subgenital plate aside. Segment 11 is similar to that of females and has a dorsal tergite (= epiproct), lateral paraprocts, and short cerci. The anus is under the epiproct and between the paraprocts. It can be seen by lifting the epiproct.
Figure 10. Male abdomen in lateral view. Orthop26L.gif

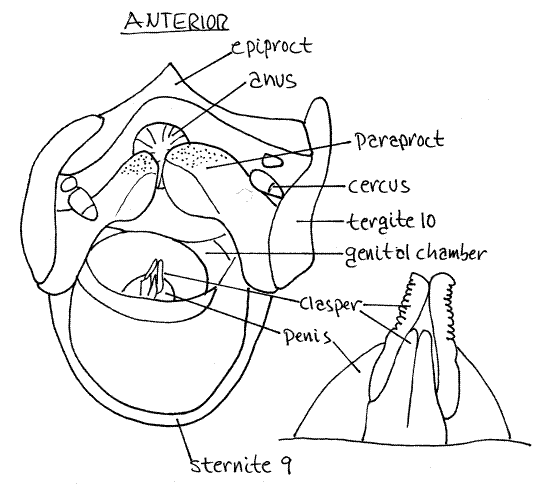
The penis consists of a soft, eversible, bulbous base bearing cuticularized valves, or claspers at its distal end (Fig 11). It contains an extension of the ejaculatory duct through which spermatophores pass during copulation. The gonopore is at the tip of the penis, between the claspers, but will not be seen. The penis lies in the genital chamber.
The genital chamber is the posterior space enclosed by the epiproct dorsally, sternite 9 ventrally and the paraprocts laterally (Fig 11). Open it by pinning the posterior tip of sternite 9 (subgenital plate) to the wax. Lift the epiproct and pin it. The space thus revealed is the genital chamber. Its floor is occupied by a large, soft, eversible penis (= aedeagus), which is an intromittent organ. Associated with the penis are dark, hard, sclerites. The male gonopore opens on the midline at the tip of the penis and in Romalea it is flanked by two pairs of slender, dark, sclerotized penis valves that hide it from view. If your specimen is a female, find a classmate with a male and ask to look at it.
Figure 11. Posterior view of the external genitalia of a male Romalea with the epiproct and sternite 9 reflected to reveal the penis. A magnified view of the tip of the penis is shown on the right. Orthop28L.gif
Internal Anatomy
Because of its large size, Romalea is well suited for the study of insect internal anatomy. The study should be conducted with the specimen immersed in fluid (tapwater, Woodring’s cricket Ringer’s solution, or 40% isopropanol) and with the aid of the dissecting microscope. Living specimens should be anesthetized or killed by placing them in a container of chloroform or ethyl acetate vapor, respectively.
" Place a living or preserved specimen in a small dissecting pan with the dorsal surface up. Push the middle legs flat against the dissecting pan and anchor them in place with an insect pin through each femur. Insert a single pin through the posterior end of the abdomen. This should be sufficient to secure the animal during the dissection. If your specimen is preserved cover it with tapwater. If it is fresh and you wish the tissues to remain alive, cover it with Woodring's cricket ringer's solution (see Supplies chapter). Use the scissors to remove the wings.
Use medium scissors to make a transverse cut across the pronotum to remove its posterior overhang. Now, slip the point of your finest scissors under the posterior edge of abdominal tergite 8, about 2 mm to one side of the dorsal midline, and cut anteriorly until you have a longitudinal dorsal incision beside the midline. Extend the cut anteriorly to the posterior edge of the truncated prothorax. by cutting through the meso- and metanota which were formerly covered by the pronotal overhand. The cut should be completely through the exoskeleton but no deeper. Avoid cutting the soft tissue under the tergites. The cut should pass dorsal to the tympanum. Make a similar incision on the other side of the animal. Extend both incisions posteriorly by inserting the scissors under the anterior edge of tergite 9 and cutting posteriorly to the epiproct.
Use fine forceps and fine scissors to separate the middorsal strip of exoskeleton from the underlying soft tissues and remove it. Try to remove the separated pieces of tergite while leaving as much soft tissue as possible behind and intact. If you are careful, the heart will remain with the body. The large body cavity (perivisceral sinus) with the gut should not be visible yet. Instead you will see only the much smaller cavity, the pericardial sinus enclosing the heart. .
During the dissection particles and debris will accumulate in the fluid of the dissecting pan and obscure your view of the specimen. When this occurs make sure everything is securely pinned to the wax of the dissecting pan and then gently pour the fluid and particles into the sink. Replace the fluid with clean and continue your study.
Hemal System
A successful dorsal dissection first reveals the heart in the pericardial sinus. These features must be destroyed to reveal the remainder of the hemocoel and its organs. Accordingly the heart must be studied now, before proceeding to the perivisceral sinus.
The arthropod body cavity is a spacious blood space, the hemocoel. In insects it is divided into three longitudinal sinuses by two horizontal diaphragms (Fig 16-7). The largest of the three is the perivisceral sinus, or perivisceral hemocoel, which contains most of the viscera. The smaller pericardial sinus lies dorsal to the perivisceral and surrounds the heart. The perivisceral and pericardial sinuses are separated by the porous dorsal diaphragm. Another small sinus, the perineural, lies ventral to the perivisceral sinus and is separated from it by the ventral diaphragm. The heart is a transparent, slender, longitudinal tube extending the entire length of the perivisceral hemocoel just ventral to the terga and dorsal to the dorsal diaphragm. It is surrounded by the pericardial sinus and blood of the hemocoel. It bears paired segmental ostia and is equipped with alary muscles to expand its lumen in diastole. Its walls contain circular muscles whose function is to constrict its lumen during systole.
Once the dorsal strip of exoskeleton is out of the way, you should see a transverse membrane, the dorsal diaphragm, which separates the relatively small, dorsal pericardial sinus from the much larger, more ventral perivisceral sinus (Fig 16-7). Both sinuses are divisions of the hemocoel and are filled with colorless blood. The dorsal diaphragm is close to the dorsal body wall and is separated from it by a space, the pericardial sinus. The diaphragm will have been destroyed if the initial scissors cuts were too deep.
The heart is an inconspicuous transparent tube on the dorsal midline of the abdomen and thorax (Fig 12, 16-7). It is closely attached to the dorsal diaphragm but is just ventral to the dorsal tergites and thus is easily lost when the exoskeletal strip is removed. Look for a narrow longitudinal tube adhering to the dorsal surface of the diaphragm exactly on the midline. Carefully tease away adhering tissue as necessary to improve your view of it. Also present in the pericardial sinus are tubular, branching, silvery tracheae, which should not be confused with the heart (Fig 14). Only the heart lies on the midline and the longitudinal tracheae are lateral to it whereas the occasional transverse tracheae cross it. Paired, segmental alary muscles extend from the heart to the body wall and diaphragm. The muscles fan out to form a thin layer over the upper surface of the dorsal diaphragm. They can be demonstrated by lifting with a minuten nadel. Individual muscles cannot be distinguished by this technique. The heart exhibits inconspicuous segmental swellings equipped with paired, segmental ostia. The swellings are apparent but the ostia will not be seen.
Figure 12. Dorsal dissection of the abdomen of a male Romalea showing the pericardial sinus and heart. Abdominal tergites are numbered. Orthop27L.gif
Perivisceral Hemocoel and Viscera
" When you have finished your study of the abdominal hemal system, perivisceral sinus, and external genitalia, remove the legs from both sides by cutting across their trochanters or coxae with medium scissors. Use fine scissors to make a middorsal longitudinal incision through the dorsal diaphragm of the abdomen. This incision should be just deep enough to cut through the thin diaphragm and should not damage the organs in the underlying perivisceral sinus. Pin the abdominal walls aside with # 4 stainless steel insect pins as you go. Note the large white air sacs of the tracheal system under the tympana in the anterior abdomen and thorax. Note also the yellow, amorphous fat body lying beneath the diaphragm throughout the hemocoel.
Upon reaching the thorax use the medium scissors to remove the heavy dorsal and lateral thoracic sclerites in pieces. Use the medium scissors to cut around a piece, then use fine scissors to cut the muscles holding the piece in place. Use heavier scissors to make the thoracic incision. Thoracic sclerites are usually too heavy to be cut by iridectomy scissors and attempting to do so will ruin them. As you cut through the thoracic sclerites, note and then cut as necessary, the massive flight muscles contained within it. Many of them originate on thoracic nota and must be cut or removed before the sclerites can be removed. Pin the walls of the thorax aside with #4 insect pins.
Without further dissection, make a preliminary inspection of the interior of the animal. The cavity occupying most of the interior is the perivisceral sinus of the hemocoel. It is part of the hemal system and is not a coelom. You will see numerous silvery, branching tubes of the tracheal system. Some of them are expanded to form air sacs. The yellow fat bodies may obscure your view of the viscera in the abdominal hemocoel. A slender, transparent, and inconspicuous, longitudinal, middorsal muscle extends the length of the abdomen just dorsal to the chief fat body. In reproductive individuals gonads will occupy much of the space in the abdomen. The regionally specialized gut extends lengthwise from the anterior mouth to the posterior anus but is presently covered by fat body and gonad so that little of it is visible. The ventral diaphragm, usually covered by yellow fat body, forms the floor of the perivisceral sinus and separates it from the perineural sinus.
Figure 13. A portion of the fat body showing its texture. Orthop29L.gif
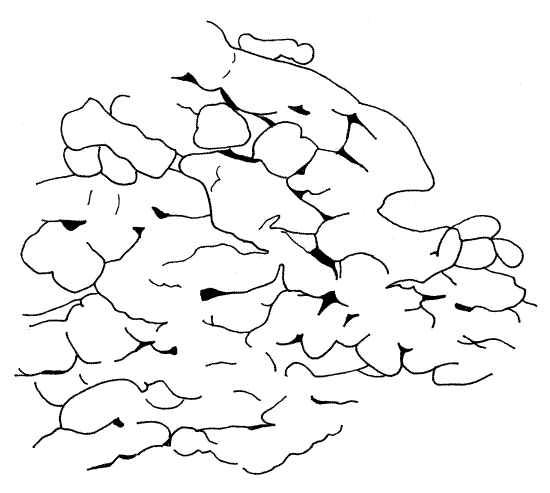
Fat Body
The fat bodies extend throughout the hemocoel but are most prevalent in the abdomen. These are sites of intermediary metabolism with a function similar to that of annelid chlorogogen tissue and the vertebrate liver. Usually they are arranged in two sheets or amorphous masses, one near the body wall and the other near the gut, and are supported by connective tissue. In Romalea the fat bodies are thin connective tissue sheets supporting small yellow lobes (Fig 13). The lobes are masses of trophocytes which are bathed in the blood of the hemocoel. The most conspicuous fat body is a sheet covering the surface of the gut in the anterior abdomen.
Respiratory System
The respiratory system, consisting of 10 pairs of spiracles (two thoracic, eight abdominal), a network of branching tubular tracheae, and several air sacs will be gradually destroyed as you study the organ systems of the perivisceral hemocoel. As you study these systems note the profusion of silvery-white tracheae extending to the organs. Each spiracle on the body surface opens into the system of interconnected tubular trachea.
Figure 14. Diagrammatic cross section of a generalized insect abdominal segment. Redrawn from Snodgrass (1935). Lepid65L.gif
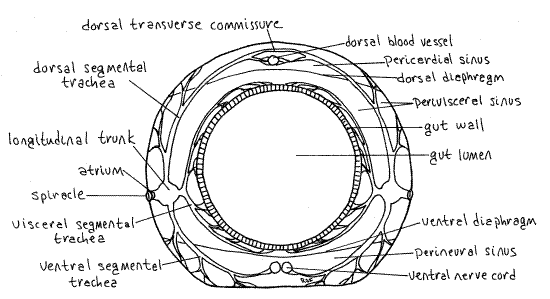
The system includes longitudinal tracheal trunks connecting the segments, branches of the trunk to the three sinuses of the hemocoel and their organs, and transverse commissures connecting right and left (Fig 13). Trachea are ectodermal invaginations of the exoskeleton and are lined by cuticle secreted by their epidermis. They branch repeatedly in a dendritic pattern and their thin walls are reinforced by spirals of chitin called taenidia (taen = band, ribbon). The number of tracheae is related to the metabolic activity of the tissues served. Air sacs are expansions of the tracheae but lack taenidia. The system is divided into cephalic, thoracic, and abdominal subdivisions.
>1a.Make a wetmount of a short piece of trachea and examine it with the compound microscope. Find the taenidia. Visualization of the taenidia will be enhanced by staining the preparation with acid fuchsin. This will stain the chitin and make it easier to see as well as confirming its composition. Let the trachea sit in the stain for 30 minutes or more to absorb the stain. <
The tracheal system includes several large, silvery, bladder-like expansions called air sacs. Their walls are not reinforced with taenidia and consequently are expansible. They function in ventilation of the tracheal system. In addition to its respiratory role, the tracheal system is important structurally and functions like connective tissue to support the tissues it pervades. The gut, as you will soon see, is a good example of an organ system supported by its tracheae.
Digestive System
Remove the fat body from the dorsal hemocoel to reveal the gut under it. Find the large gonad covering the posterior regions of the gut in the posterior hemocoel (Fig 15). Remove the fat body covering the gonad then use fine scissors to free it (the gonad) from its anterior connections with the remaining viscera. Note that most of these connections are tracheae. Locate the gonoducts (oviduct or sperm duct) exiting the gonad laterally. Cut the duct on the left so the gonad can be reflected to the right, thereby revealing the posterior gut but keeping the gonad intact for later study.
Figure 15. Dorsal dissection of a male Romalea showing the digestive system. The dorsal diaphragm and heart have been removed, most of the Malpighian tubules and most of the epiproct has been removed. The few Malpighian tubules in the drawing are shown much shorter and much less numerous than in life. The testis has been removed but its position is indicated by a dashed line. The ovary occupies a similar position in females. Orthop30L.gif
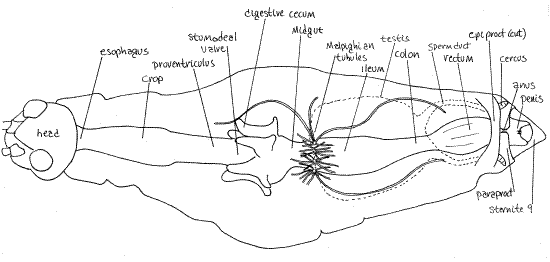
Like that of other arthropods, the gut consists of anterior ectodermal foregut, middle endodermal midgut, and posterior ectodermal hindgut. The foregut consists of mouth, pharynx, esophagus, crop, and proventriculus and is lined by a cuticle secreted by its epidermal epithelium. The gut begins, of course, with the mouth which opens from the preoral cavity but it cannot be seen from your present vantage point inside the hemocoel. The mouth opens into the short, narrow pharynx, which is entirely contained within the head capsule and consequently is likewise not visible at present.
The pharynx leads to the esophagus, which is largely confined to the head but extends a short distance through the foramen magnum to enter the anterior prothorax. (Fig 15, 21-8A). Posterior to the esophagus the gut widens to become the large crop. The separation between esophagus and crop is not distinct on the exterior but will be obvious later when you open the gut. Next, the gut narrows again as it becomes the proventriculus, or gizzard.
The proventriculus, which is the last region of the foregut, ends where the proventriculus joins the short midgut. An elaborate stomodeal valve, visible only from inside the gut, marks the separation of the fore- and midguts. Protruding conspicuously from the periphery of the midgut are six hollow diverticula, the digestive cecae.
The remainder of the gut is hindgut, which accounts for about half the length of the gut. The foregut accounts for most of the remaining half with the midgut being short. Internally the proctodeal valve marks the midgut-hindgut junction. The junction of midgut and hindgut is marked externally by clusters of long slender Malpighian tubules (Fig 15). These form a tight mass in the vicinity of the junction but also extend anteriorly into the thorax and posteriorly to the posterior end of the abdomen. The hindgut, also known as the intestine, consists of three regions. First is the large ileum followed by the short, narrow colon. The longitudinally ridged rectum is the final region and it opens via the anus to the exterior. Like the foregut, the hindgut is lined by epidermis, which secretes a cuticle.
" Some gut regions are difficult to distinguish from adjacent regions from the exterior whereas the epithelia lining their lumina are distinct. Use fine scissors to open the entire length of the gut from esophagus to anus with a longitudinal incision.
The interior of the esophagus can be seen as it emerges from the head capsule. From the inside, its walls are longitudinally folded whereas those of the crop bear fine oblique ridges best seen at 40X. Each ridge bears a row of tiny, dark, cuticular denticles (= little teeth) which can be seen by focusing carefully with 30-40X. The floor of the crop features a wide, midventral, longitudinal typhlosole composed of a pair of parallel longitudinal ridges flanking a median groove. Observe the typhlosole at 10X. The epithelium lining the proventriculus is longitudinally ridged and the ridges bear tiny conical denticles similar to those of the crop. Find the longitudinal ridges and their denticles by focusing carefully with 40X.
Using 10X again, observe the interior of the stomodeal valve at the foregut-midgut junction. The final region of the foregut (proventriculus) is recognized by its numerous, fine, longitudinal ridges whereas the beginning of the short midgut can be identified by its 5-6 wide transverse ridges. Between the two is the conspicuous stomodeal valve marked by six wide, but short, longitudinal ridges.
The openings of the digestive cecae are immediately posterior to the ridges of the stomodeal valve and anterior to the transverse ridges. Find at least one and probe it with a minuten nadel to demonstrate that it is hollow and that its lumen is continuous with that of the midgut. The midgut is divided into two regions whose epithelia differ. The anterior midgut bears the wide transverse ridges just mentioned whereas the posterior midgut, which is about the same length, has its walls thrown into inconspicuous longitudinal folds. The longitudinal folds end with the proctodeal valve which marks the end of the midgut and beginning of the hindgut. The openings to the Malpighian tubules are located at the midgut-hindgut junction but will not be seen.
Notice that the contents of the hindgut are enclosed in a transparent peritrophic membrane. The walls of the ileum are thin and have inconspicuous longitudinal ridges similar to those of the posterior midgut. These folds become better organized in the colon where they are clearly visible as six distinctive ridges. In the rectum the six ridges remain but become wide and flat-topped. These six flat white bands of cells are known as rectal papillae.
The salivary glands will not be seen until you begin dissection of the ventral nerve cord. They lie under the ventral diaphragm and have not yet been exposed.
Excretory System
The chief insect excretory organs are the Malpighian tubules, which were seen earlier arising at the midgut-hindgut junction (Fig 15, 21-8A). These are long, slender, hollow diverticula of the gut lumen. In Romalea the tubules are very numerous and very long, extending anteriorly and posteriorly from the midgut-hindgut junction. They extend into the perivisceral hemocoel where they are immersed in blood. Nitrogenous breakdown products of protein metabolism, mostly ammonia, are released into the hemocoel, absorbed by the trophocytes of the fat body, converted to uric acid, and returned to the blood. The uric acid is them absorbed by the tubules, precipitated, and released into the hindgut where it becomes part of the feces.
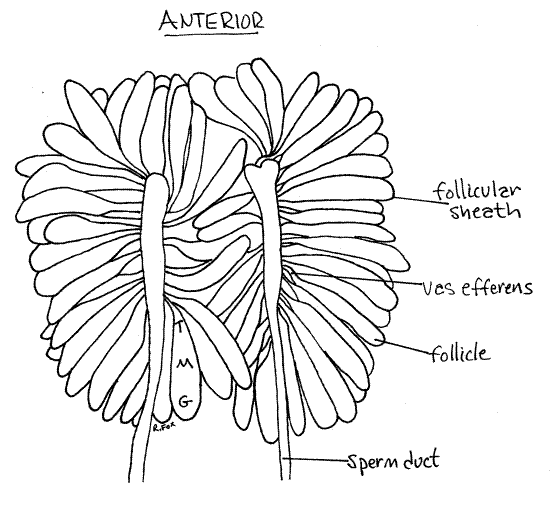
Figure 16. The testes of a male Romalea in ventral view. G = germarium, M = zone of maturation, T = zone of transformation. Orthop31L.gif
Reproductive System
Study both male and female reproductive systems by sharing specimens with a classmate. If you have followed instructions, the gonad will be displaced to the right but will be intact and available for study.
Male
The male system begins with a pair of dorsal testes (Fig 15, 16, 21.12A). The two testes are coalesced over the dorsal midline and appear to be a single testis. A sperm duct (=vas deferens) exits each posterior lateral corner of each testis. The ducts are wrapped in fat body and are easy to miss. The left one was cut earlier but should still be present and recognizable. Find it. Each duct extends laterally and posterior to disappear under the gut. Reflect the testis, without cutting the remaining sperm duct, posteriorly to reveal its ventral surface (Fig 16).
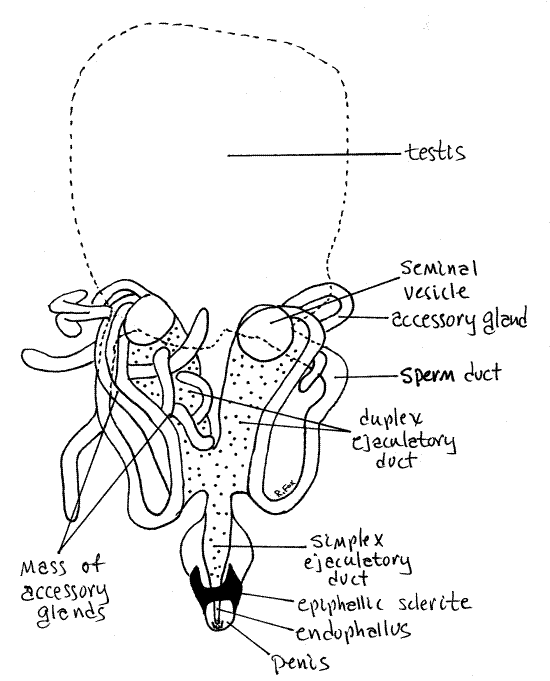
Fig 17. Dorsal view of the reproductive system of a male Romalea. For clarity, the testis is shown as a dashed line. Most of the accessory glands have been omitted. The ejaculatory duct is stippled. Orthop32L.gif
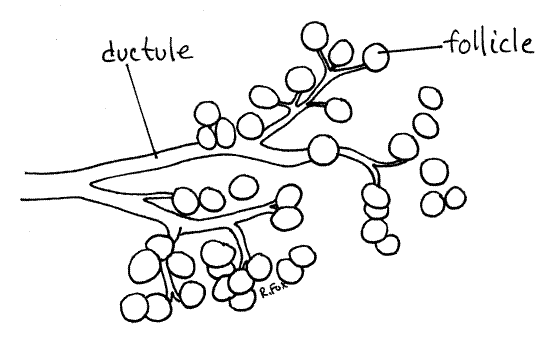
From the ventral surface each testis can be seen to consist of numerous elongate follicles (= sperm tubes) draining into a central white sperm duct (= vas deferens) (Fig 16). Each follicle is a blind ending peritoneal sac lined with epithelium and connecting with the sperm duct by a narrow vas efferens.
Spermatogenesis occurs in the follicles beginning at the blind, upstream end where diploid spermatogonia are located in a zone known as the germarium (Fig 16). As developing cells move down the follicle toward the vas efferens and sperm duct, they undergo the successive divisions of gametogenesis and by the time they reach the vas efferens they are spermatozoa. The follicle is divisible into consecutive zones which are, from upstream to downstream, the germarium with spermatogonia, zone of maturation with primary and secondary spermatocytes, and zone of transformation where spermatids become spermatozoa.
Each sperm duct curves ventrally around the gut to join the ejaculatory duct which leads to the penis and gonopore (Fig 17). Cut across the gut in the middle of the ileum, free the posterior gut from the body wall, and pin it aside to reveal the ventral regions of the reproductive system. Trace the sperm ducts around the rectum to the ejaculatory duct. Upstream the ejaculatory duct is double (duplex ejaculatory duct) but the two upper arms join each other distally to form a single duct (simplex ejaculatory duct) that enters the penis. Associated with each duplex ejaculatory duct is a bright white seminal vesicle for sperm storage. Numerous blind ending, tubular accessory glands extend from the duplex ejaculatory ducts and form a loose mass around the duct. Some of the accessory glands are bright white and some are transparent. Some are long and some short. Some extend away from the mass and others are confined to it.
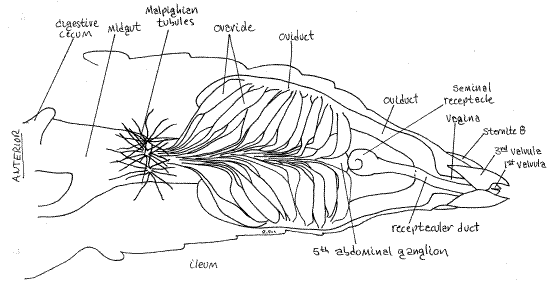
Figure 18. Dorsal view of the reproductive system of a female Romalea. Malpighian tubules are drawn much shorter and less numerous than in life. Orthop38L.gif
Female
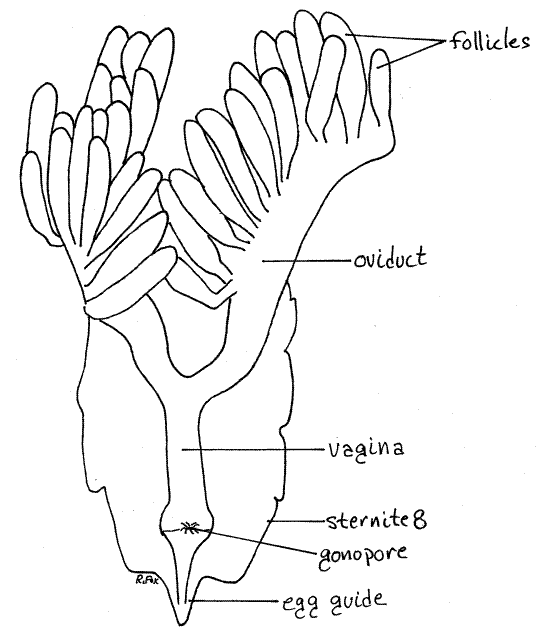
The female reproductive system consists of two ovaries, two oviducts and a common vagina opening by the gonopore between the two first valvulae (Fig 9, 18, 22, 21-11). In addition, an independent seminal receptacle opens to the exterior via a separate receptacular duct and receptacular pore. Sperm stored in the seminal receptacle are used to fertilize the eggs during oviposition.
The two ovaries are the most conspicuous feature of the female system because they are large, bright yellow, and dorsal most. They are located in the posterior abdomen, dorsal to the gut and ventral to the dorsal diaphragm. You need remove only the investing layer of fat body and tracheae to reveal them. Each is a cluster of tubular ovarioles, similar to the testicular follicles, emptying into a lateral gonoduct. The two ovaries of reproductively mature females are large and occupy, along with the fat body, most of the space in the dorsal posterior abdomen, between the dorsal diaphragm and the gut (Fig 15, 22).
Mature eggs in the ovaries are bright yellow and, since they account for most of the volume of the ovary, impart their color to these organs. Immature eggs are white. The fat body investing the ovaries is also yellow and care must be taken that the two are not confused. Remove as much of the fat body as possible from the ovary. Each ovary is composed of numerous tubes, known as ovarioles (Fig 18, 19), homologous to the follicles, or sperm tubes, of the testis. In each ovary a series of ovarioles drain into a lateral oviduct. Each ovary has an oviduct extending along its outside border.
The ovarioles begin upstream as tiny filamentous tubes that gradually increase in diameter moving downstream (Fig 19). At the downstream end the ovariole exhibits its maximum diameter, after which it narrows before joining its oviduct. Like the follicles, each ovariole consists of a germarium of gonial cells (oogonia) at the narrow upstream end. Daughter cells produced by mitotic divisions of oogonial cells become either more gonial cells or primary oocytes. The oocytes move downstream in the tube and undergo meiosis I to become secondary oocytes. These also move downstream and become large, oval, yellow "eggs" in the distal end of the ovariole. During oviposition these eggs exit the ovariole and pass through the oviduct to the vagina where they are fertilized by sperm from the seminal receptacle. Following fertilization the eggs are inserted into the ground by the valvulae of the ovipositor. The final meiotic division occurs after fertilization. The pronuclei unite and a zygote is formed.
Fig 19. A single ovariole from a Romalea ovary. Orthop41L.gif
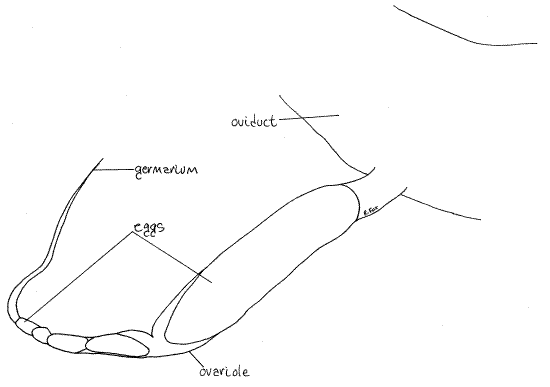
The two oviducts depart from the posterior lateral corner of their respective ovaries and curve far ventrally, around the gut, to coalesce on the midline ventral to the gut and form the single vagina. Observation of the vagina must be postponed, however, in order to find and study the seminal receptacle which is dorsal to it.
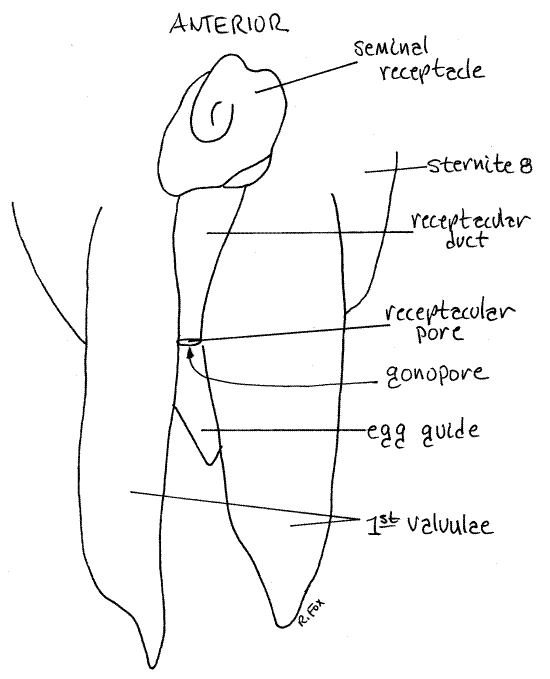
" Cut across the posterior rectum where it joins the anus and pull the intestine forward, between the two ovaries and out of the field of view. This will reveal the last ganglion of the ventral nerve cord and the seminal receptacle. The receptacle is nestled against the curved posterior border of this ganglion (Fig 18, Fig I, Fig 9). Try to avoid damaging the ganglion as the nervous system has not yet been studied. Remove the third valvulae (Fig 20). The receptacular duct extends to the receptacular pore between the two first valvulae to the spermatheca. The duct is coiled around the vesicle but that is not apparent without careful dissection.
The receptacular pore is near the gonopore (Fig 21, 20). The gonopore is at the base of a fingerlike, sclerotized process on sternite 8, the egg guide, located ventrally in the genital chamber (Fig 9). The spermathecal pore is a little dorsal to it.
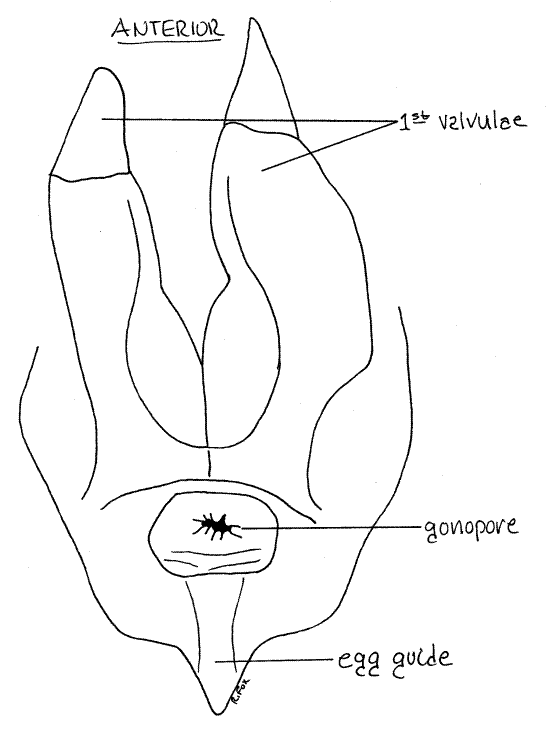
Remove the seminal receptacle, its duct, and the first valvulae. Trace the lateral oviducts from the ovaries posteriorly toward the midline where they join to form the median vagina (Fig 18, 19, 22, 21-11A). The vagina is ventral to all other organs in the posterior hemocoel, including the nervous system. Find the gonopore at the base of the egg guide.
Figure 20. Dorsal view of the posterior end of a female Romalea. The epiproct, paraprocts, and third valvulae have been removed and the first valvulae are in their normal position, thus revealing their dorsal surface. With the valvulae in this position the receptacular pore is exposed to view. The gonopore is ventral to the receptacular pore, between it and the egg guide. Orthop40L.gif
Nervous System
The nervous system is divisible into the central nervous system (CNS) consisting of the brain and ventral nerve cord, and the peripheral nervous system (PNS) consisting of sensory and motor neurons connecting the CNS with the tissues. The PNS has somatic and visceral components but will not be studied in this exercise. The central nervous system consists of a ventral nerve cord in the thorax and abdomen and a dorsal brain in the head. If you are working with fresh, unpreserved material, the nerve cord may be difficult to see against a similar background. Be sure the specimen is firmly pinned to the dissecting wax and then gently pour the water out of the pan. Replace it with 40% isopropanol. The alcohol will render the nerve cord opaque, white, and easier to see. This effect improves with time.
Figure 21. Dorsal view of the posterior end of a female Romalea. The epiproct, paraprocts, and third valvulae have been removed and the first valvulae reflected dorsally to reveal their ventral surface. This view exposes the gonopore. Orthop39L.gif
Ventral Nerve Cord
" Remove the reproductive system and discard it. Cut across the esophagus and remove the gut so you can see the floor of the perivisceral sinus. The floor is the ventral diaphragm. Through it you can see the perineural sinus and the double ventral nerve cord. The nerve cord is a white band immediately inside the diaphragm. The nerve cord adheres to the ventral surface of the ventral diaphragm. The fat body covers its dorsal surface. Carefully remove the ventral diaphragm and fat body from the nerve cord over the entire length of the abdomen and thorax. In the thorax the massive flight muscles must also be removed. The abdominal region of the cord is easily revealed but considerably more digging is required to find the thoracic region. Remove all tissue necessary to reveal the entire length of the ventral nerve cord.
Figure 22. Dorsal view of the female reproductive system. The gut, seminal receptacle, epiproct, paraprocts, and valvulae have been removed to reveal the vagina. Orthop42L.gif
In the thorax removal of the fat body and ventral diaphragm will reveal the salivary glands, of which there are two. Each gland consists of numerous white spherical follicles arranged in clusters, rather like clusters of grapes (Fig 23). The clusters are connected by branches of the two salivary ducts. The ducts extend anteriorly into the head and coalesce to form a single duct which empties into the salivarium of the preoral cavity. Where the salivary glands obscure the ventral nerve cord they must be removed.
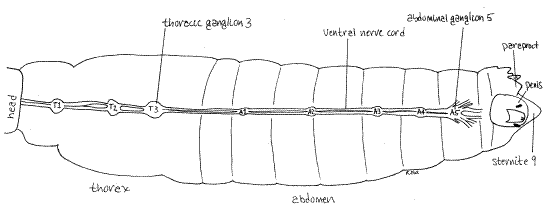
The ventral nerve cord is conspicuously double and connects a chain of ganglia on the floor of the perineural sinus (Fig 24). The ganglia consist of three large thoracic ganglia, T1-T3 and five smaller abdominal ganglia, A1-A5. Notice the abundant nerves entering and leaving the thoracic ganglia in contrast with the relatively few of the abdominal ganglia. Remember that the thorax is the location of three pairs of legs and two pairs of wings and houses the myriad muscles that operate them.
Figure 23. A small portion of a salivary gland. Orthop33L.gif
Brain
" Dissection of the head capsule to reveal the brain is an optional exercise. The brain is well protected in the heavily sclerotized cranium and exposing can be difficult. Use medium scissors and forceps to open the cranium. Do not attempt to cut or manipulate the hard cranium with your delicate iridectomy scissors or microdissecting forceps. It will ruin them.
Figure 24. Dorsal dissection of the postcranial nervous system of a male Romalea. Heart, fat bodies, dorsal and ventral diaphragms, and gut have been removed. Orthop34L.gif
Cut across the esophagus and remove it. Cut across the posterior cervix to separate the head from the body. Look into the severed cervix and find the foramen magnum. This is the best view of it you have had. Find the severed ventral nerve cord where it enters the foramen magnum. Find the severed esophagus. Notice the abundant muscles filling the head capsule. These are responsible for operating the head appendages, especially the mandibles. The brain is dorsal to the esophagus and is connected to the ventral nerve cord by a pair of circumesophageal connectives.
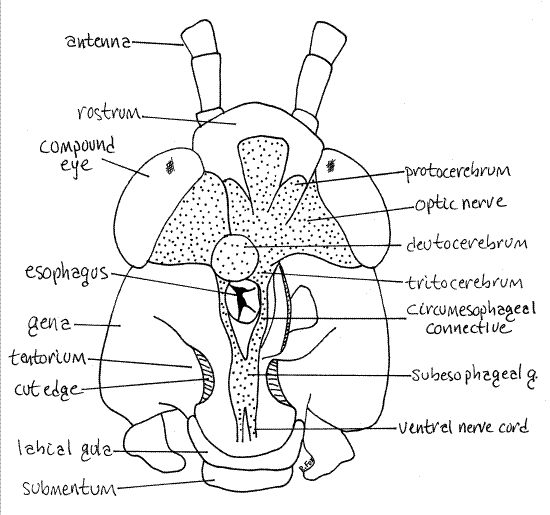
Figure 25. Dorsal dissection of the head capsule of Romalea with the vertex and mandibular muscles removed to reveal the brain. The right deutocerebrum is omitted for clarity. The esophagus is cut transversely. The nervous system is stippled. Orthop35L.gif
With medium scissors make two longitudinal cuts anteriorly through the cranium from the foramen magnum, one on each side of the vertex. Make the cuts as shallow as possible to avoid damage to the brain. Extend these incisions anteriorly dorsal to the eyes to the frons. Remove the strip of cuticle thus freed. The space opened if filled with muscles and the esophagus. Insert a #4 insect pin through the labium into the wax to hold the head in place. Cut away the posterior portion of the genae. Use fine forceps to remove the muscles in the posterior head. Do not remove the esophagus, brain, or nerve cord. The muscle has a fibrous texture lacking in nerve tissue. Nerve tissue is yellowish, muscle is dull white. Most of the muscles extend from the cranium to two pairs of thin, flat, transparent mandibular apodemes (Fig 25). The medial apodemes is the larger and extends dorsally almost to the vertex. The later apodeme is smaller.
As the muscles are removed the brain will gradually come into view (Fig 25). It consists of the three major paired ganglia of the mandibulate brain which make up the three brain regions. These are the protocerebrum, deutocerebrum, and tritocerebrum, of which the deutocerebrum is dorsalmost. A large optic nerve extends laterally from each protocerebrum to the nearby compound eyes. A pair of circumesophageal connectives extend from the tritocerebrum around the esophagus to unite at the subesophageal ganglion ventral to the esophagus. The paired ventral nerve cord extends posteriorly from the subesophageal ganglion. Remove the soft tissue of the cervix, including its cuticle. Remove all non-nervous tissue surrounding the esophagus, including the medial mandibular apodeme. This will expose a heavy transverse ventral bar of cuticle just ventral to the esophagus. This is the central body of the tentorium, the all-important internal structural brace of the head capsule. The ventral nerve cord enters the head capsule ventral to the tentorium. The subesophageal ganglion is immediately dorsal to the central body of the tentorium. The latter must be removed to reveal the ganglion and the circumesophageal connectives. The subesophageal ganglion represents the combined ganglia of the mandibles, maxillae, and labium.
References
Albrecht FO . 1953. The Anatomy of the Migratory Locust. Althone Press, London. 118p.
Brooks WK . 1890. Handbook of Invertebrate Zoology. Bradlee Whidden, Boston. 352p.
Capinera JL, Scott RD, Walker TJ. 2004. Field guide to grasshoppers, katydids, and crickets of the United States. Comstock Pub. Assoc, Cornell Univ. Press, Ithaca. 249pp.
Chapman RF. 1998. The insects, 4 th ed. Cambridge Univ. Press, Cambridge. 770pp.
Comstock JH, Kellog VL . 1916. The Elements of Insect Anatomy. Comstock, Ithaca. 145p.
Eisner T. 2003. For love of insects. Belknap Press, Harvard Univ. Press, Cambridge. 448 pp.
Gilbert C. Entomology, 2 nd ed. Plenum Press, New York.
Harrison FW, Locke M. 1998. Microscopic anatomy of invertebrates, 11A. Insecta. Wiley-Liss, New York. 1-381pp.
Harrison FW, Locke M. 1998. Microscopic anatomy of invertebrates, 11B. Insecta. Wiley-Liss, New York. 385-840 pp.
Harrison FW, Locke M. 1998. Microscopic anatomy of invertebrates, 11C. Insecta. Wiley-Liss, New York. 843 -1296 pp.
Ruppert EE, Fox RS, Barnes RB. 2004. Invertebrate Zoology, A functional evolutionary approach, 7 th ed. Brooks Cole Thomson, Belmont CA. 963 pp.
Snodgrass RE . 1935. Principles of Insect Morphology. McGraw-Hill, New York. 667p.
Snodgrass RE . 1952. A Textbook of Arthropod Anatomy. Cornell, Ithaca. 363p.
Whitman D. 2006. Personal communication.
Supplies
Dissecting microscope
Small dissecting pan or dish (a sardine tin with a wax bottom is ideal)
Living anesthetized, freshly sacrificed, or preserved Romalea
Carbon dioxide, chloroform, or ethyl acetate for living specimens
Woodring’s cricket Ringer’s solution
40% isopropanol