Invertebrate Anatomy OnLine
Pachydesmus ©
Flat-backed Millipede
29jun2006
Copyright 2005 by
Richard Fox
Lander University
Preface
This is an exercise from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises can be accessed by clicking on the links on the left. A glossary and chapters on supplies and laboratory techniques are also available. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Panarthropoda SP, Arthropoda P, Myriapoda SC, Progoneata, Dignatha, Diplopoda C, Chilognatha, Helminthomorpha, Eugnatha, Merochaeta SO, Polydesmida O, Xystodesmidae F (Fig 6-15, 20-14, 20-15)
Panarthropoda SP
Panarthropoda includes Onychophora, Tardigrada, and Arthropoda. These taxa share segmentation, a hemocoel, saccate nephridia, ecdysis of a secreted chitinous but non-collagenous exoskeleton, loss of locomotory cilia, a tubular, dorsal, ostiate heart in a pericardial sinus, a coelom reduced to end sacs and gonocoel, and paired segmental legs.
Arthropoda
Arthropoda, by far the largest and most diverse animal taxon, includes chelicerates, insects, myriapods, and crustaceans as well as many extinct taxa. The body is segmented and primitively bears a pair of jointed appendages on each segment. The epidermis secretes a complex cuticular exoskeleton which must be molted to permit increase in size. Extant arthropods exhibit regional specialization in the structure and function of segments and appendages. The body is typically divided into a head and trunk, of which the trunk is often itself divided into thorax and abdomen.
The gut consists of foregut, midgut, and hindgut and extends the length of the body from anterior mouth to posterior anus. Foregut and hindgut are epidermal invaginations, being derived from the embryonic stomodeum and proctodeum respectively, and are lined by cuticle, as are all epidermal surfaces. The midgut is endodermal and is responsible for most enzyme secretion, hydrolysis, and absorption.
The coelom is reduced to small spaces associated with the gonads and kidney. The functional body cavity is a spacious hemocoel divided by a horizontal diaphragm into a dorsal pericardial sinus and a much larger perivisceral sinus. Sometimes there is a small ventral perineural sinus surrounding the ventral nerve cord.
The hemal system includes a dorsal, contractile, tubular, ostiate heart that pumps blood to and from the hemocoel. Excretory organs vary with taxon and include Malpighian tubules, saccate nephridia, and nephrocytes. Respiratory organs also vary with taxon and include many types of gills, book lungs, and tracheae.
The nervous system consists of a dorsal, anterior brain of two or three pairs of ganglia, circumenteric connectives, and a paired ventral nerve cord with segmental ganglia and segmental peripheral nerves. Various degrees of condensation and cephalization are found in different taxa.
Development is derived with centrolecithal eggs and superficial cleavage. There is frequently a larva although development is direct in many. Juveniles pass through a series of instars separated by molts until reaching the adult size and reproductive condition. At this time molting and growth may cease or continue, depending on taxon.
Mandibulata
Mandibulata includes arthropods in which the third head segment bears a pair of mandibles. As currently conceived this taxon includes myriapods, hexapods, and crustaceans. Appendages may be uni- or biramous and habitats include marine, freshwater, terrestrial, and aerial.
Tracheata
Myriapods and hexapods share tracheae and a single pair of antennae and are sister taxa in Tracheata. Crustaceans, which have gills and lack tracheae, are excluded and form the sister group.
Myriapoda
The body consists of a head and trunk with numerous segments but no tagmosis into thorax and abdomen. Myriapoda includes the familiar centipedes and millipedes, as well as symphylans and pauropods.
Dignatha
This myriapod taxon comprises millipedes and pauropods, both of which have two pairs of mouthparts instead of the three of the ancestral tracheate, insects, centipedes, and symphylans. Millipedes have a pair of mandibles and a single gnathochilarium, which probably arose through fusion of the two first maxillae. The second maxillae are lost in dignathans.
Diplopoda
Most millipedes are elongate, often worm-shaped, mandibulates, although some are short and resemble woodlice. The body consists of a head and a trunk of many segments, most of which bear paired appendages. Most of the trunk segments occur in fused pairs, called diplosegments, each of which bears two pairs of legs and two pairs of spiracles externally. Internally each diplosegment has two pairs of ganglia, two pairs of ostia and two pairs of lateral blood arteries.
The head bears a pair of antennae, a pair of mandibles, and a pair of fused first maxillae. There is no second maxilla (= labium) and its segment is absent. The exoskeleton is strengthened with calcium salts and is correspondingly harder than that of most hexapods. The heart is an elongate ostiate dorsal tube extending most of the length of the body. Excretion is accomplished via a single pair of Malpighian tubules and a pair of maxillary glands (= saccate nephridia). Respiration is via segmentally arranged tracheae with segmental spiracles.
The nervous system consists of a three-part mandibulate brain and a double, ganglionated, ventral longitudinal nerve cord. The subesophageal ganglion consists solely of the mandibular and maxillary ganglia. Millipedes are gonochoric with the gonopores located anteriorly, on segment 3. Sperm transfer is indirect with intromittent organs and spermatophores. Eggs are laid in the summer and most species overwinter as adults. Juveniles hatch with three pairs of legs and add body segments and additional legs with successive molts (Fig 20-12F). Sexual dimorphism is slight.
Many, including polydesmids, are chemically protected by repugnatorial (scent) glands that produce and release toxic or repellant substances including aldehydes, phenol, quinones, iodine, hydrogen cyanide, and chlorine.
Figure 1. Dorsal view of the trunk of a female polydesmid millipede. The head is hidden by the first trunk segment. Milliped22L.gif
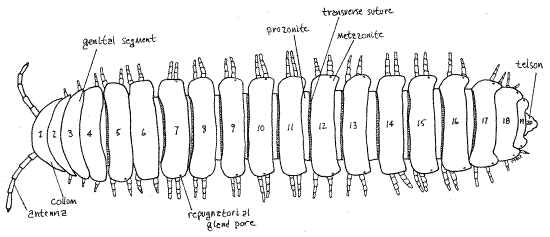
Polydesmida O
Polydesmids, the flat-backed millipedes, are relatively large (up to 13 cm but with some as small as 3 mm) millipedes with 20 rings ("segments”). Polydesmids range in size from 0.3 to 13 cm. Pachydesmus contains the largest North American Polydesmida and some are common and familiar animals of lawn and garden. The body proper, although circular in cross section, appears to be, and functionally is, dorsoventrally flattened due to large lateral plates (paranota) which extend from the sides of the cylindrical body (Fig 1, 8, 20-8A, 20-10C). The four exoskeletal sclerites of each segment and diplosegment are fused together to form a rigid ring. Many are brightly (aposematically) colored, frequently with a dark, often black, background with contrasting bright red, orange, yellow, or cream markings. Polydesmids are blind without ocelli or eyes. This is the largest millipede order with more than 2700 known species (as of 1992).
Natural History
Millipedes, in general, are negatively phototactic, nocturnal detritivores that feed on dead vegetation. A few eat living plants and can be a problem in greenhouses but most consume decaying leaves and wood. A few are carnivores feeding on other arthropods and earthworms. Millipedes live in moist habitats including soil, leaf litter, under stones, decaying logs, under bark, or in caves. Millipedes join earthworms in assuming the chief responsibility for recycling organic molecules in forest ecosystems. About 10,000 species have been described but tens of thousands more are thought to await discovery, especially in the tropics, where millipedes are most diverse. Five higher taxa (superorders) are recognized.
Polydesmids are dorsoventrally depressed millipedes belonging to an eco-morphological group known as “flat wedgers”. Wedgers live in the spaces between flat surfaces such as layers of decaying leaves on the forest floor. The flat shape and powerful musculature allows the animal to push its way between the leaves, wedging them apart.
Figure 2. Ventral view of the head and anterior trunk. The legs have been omitted from ring 6 to reveal the two spiracular apodemes. Milliped23L.gif
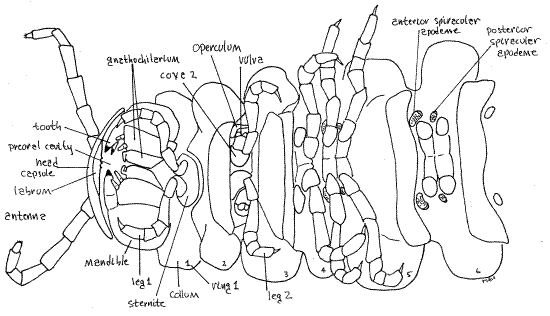
Laboratory Specimens
This exercise was written using specimens of Pachydesmus collected in Greenwood Co. South Carolina but it can be used with any polydesmid millipede. Living, freshly sacrificed, or preserved specimens can be used. Anesthetization can be accomplished with chloroform or carbon dioxide. Living specimens will release an odiferous repugnatorial substance when handled. The study should be conducted with magnification, preferably that of a dissecting microscope. This exercise is limited to external anatomy.
External Anatomy
Exoskeleton
The millipede body, like that of all arthropods, is covered by a secreted exoskeleton, or cuticle, composed of chitin and protein. In some arthropods, notably millipedes and crustaceans, the cuticle is hardened, not only by sclerotization (= protein tanning) but also by the incorporation of calcium salts. Consequently millipede sclerites are often very hard, rocklike, and totally inflexible. Millipedes play an important role in calcium cycling in many terrestrial ecosystems.
> a. If an extra specimen (dead) is available for the purpose, remove a segment from its body and place it in a small culture dish of 10% hydrochloric acid. With fine forceps and needles test the rigidity of the cuticle and note its ability to resist deformation. Cover the dish and set it aside, looking at it occasionally during the laboratory period. You will notice the slow evolution of small bubbles of carbon dioxide resulting from the reaction between the acid and calcium carbonate in the cuticle. At the end of the period test the rigidity again, trying to bend the cuticle with your forceps. Is there any difference? Try again after 24 hours or during the next lab period. Now is there a difference? How do you account for the flexibility of the cuticle after a day or two in acid? <
Figure 3. Left side view of the head. Milliped25L.gif
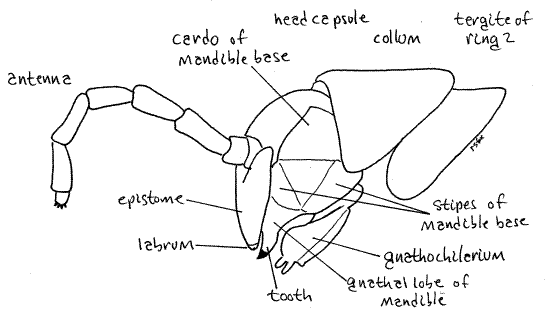
Tagmata
The elongate body is divided into two distinct tagmata, the short anterior head and the long, obviously segmented, posterior trunk (Fig 1, 20-9A).
Head
The head consists of a smooth sclerotized head capsule (= cranium, epicranium) to which is fused the labrum and with which the three pairs of head appendages articulate (Fig 3). The capsule forms the dorsal and anterior surfaces of the head. The unusually large mandibles are the sides and the gnathochilarium (= lower lip) is the ventral surface. Posteriorly the head joins the trunk via a short neck. The region of the head capsule ventral to the antennae is the epistome (Fig 3, 4). The small, narrow, toothed labrum (= upper lip) is immovably fused to the epistome ventrally (Fig 4, 6). Anteriorly the head capsule is deeply invaginated to form the dorsal wall of the preoral cavity (more later).
Antennae
The anteriormost head appendages are the two filiform antennae, each consisting of 7 cylindrical articles (Fig 3, 4, 20-9). The antennae articulate with the anterior/dorsal surface of the head capsule. The tip of the distal antennal article has four tiny chemoreceptive cones characteristic of diplopods, as well as numerous mechanoreceptive setae. A walking millipede continuously taps the ground with the tips of its antennae.
Figure 4. En face view of the head. Milliped28L.gif
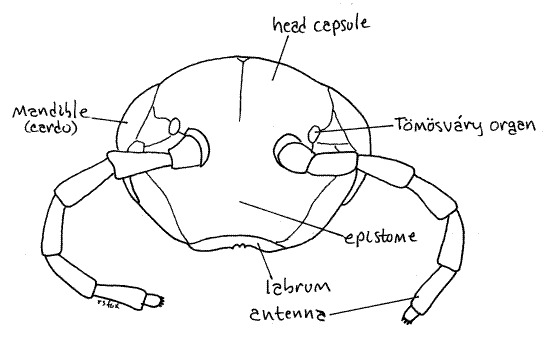
Mouthparts
Diplopods and pauropods differ from other tracheates (chilopods, symphylans, and insects) in having only two pairs of mouthparts (mandibles, gnathochilarium) instead of the usual three (mandibles, maxillae, labium). The former form a taxa known as Dignatha, in reference to their two pairs of mouthparts (di = two, gnath = jaw) whereas the latter are trignathan. The dignathan mandibles, of course, are homologous to those of other tracheates but the origin of the gnathochilarium is disputed. Some believe it to be homologous to the fused first maxillae (= maxillae) of tracheates that have lost the labium (= second maxillae) whereas others think it arose through the combination of the maxilla and labium of the ancestral tracheate.
Gnathochilarium
The labium-like gnathochilarium (= lower lip) is the third pair of head appendages and is posterior to the mandibles. It is best studied before the mandibles, however, because it almost completely covers and hides the jaws of the mandibles from view. Like the insect labium, the gnathochilarium is a single piece formed in development by the median fusion of two lateral appendages. It is a broad flat plate ventral on the millipede head (Fig 1, 5, 20-10A).
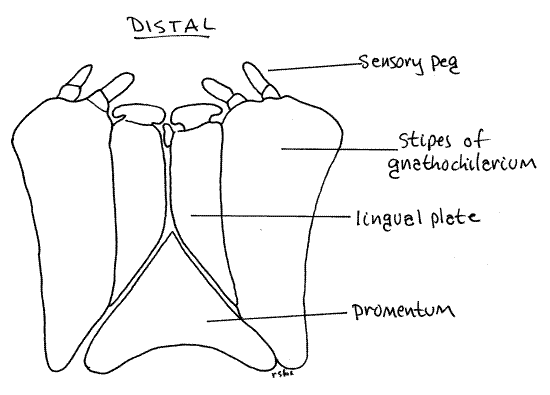
The gnathochilarium consists of two large lateral stipes, a median promentum (= mentum, prementum), and two medial lingual plates (= lamellae linguales) (Fig 5). Each stipes has two distal sensory pegs bearing contact chemoreceptors (taste).
Figure 5. Posterior-ventral view of the gnathochilarium as seen in an intact, undissected animal. Milliped29L.gif
Mandibles
The mandibles are the second pair of head appendages and first pair of mouthparts. Millipede mandibles are large, powerful and heavy, making up most of the side of the head (Fig 3). Each consists of a heavy, immovable, biarticulate mandibular base and a toothed, movable, distal gnathal lobe. The mandibular base is best viewed from the side (Fig 3) where it forms most of the side of the head. It consists of a proximal basal article, the mandibular cardo, and a distal basal article, the mandibular stipes. The gnathal lobe is also visible laterally but to see it in more detail use forceps to pull the free edge of the gnathochilarium posteriorly and look at from the ventral aspect. The gnathal lobe has two heavily sclerotized movable teeth, the outer tooth and the inner tooth, a rasp-like molar plate, and a set of pectinate lamellae (Fig 6). The teeth are conspicuous in an intact, undissected specimen, the pectinate lamellae are exposed to view but not conspicuous, and the molar plate is deep in the preoral cavity and difficult to see without further dissection.
Figure 6. The right mandible in ventral view with the gnathochilarium and left mandible removed. Milliped31L.gif
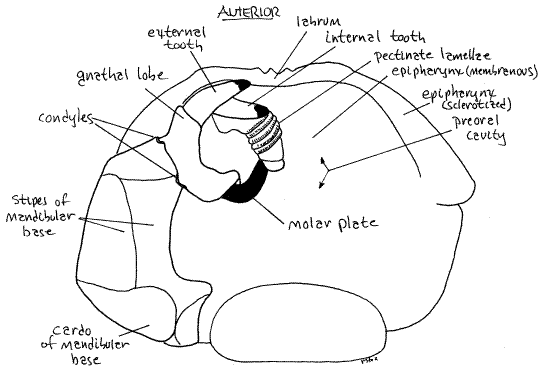
The gnathal lobes articulate with the stipes of the base via two condyles (Fig 6) and, during feeding, are moved transversely to oppose each other. The teeth are for cutting and the molar is for crushing or grinding. The first three pairs of legs may function as auxiliary mouthparts to manipulate food items and transfer them to the jaws (reminiscent of the maxillipeds of crustaceans).
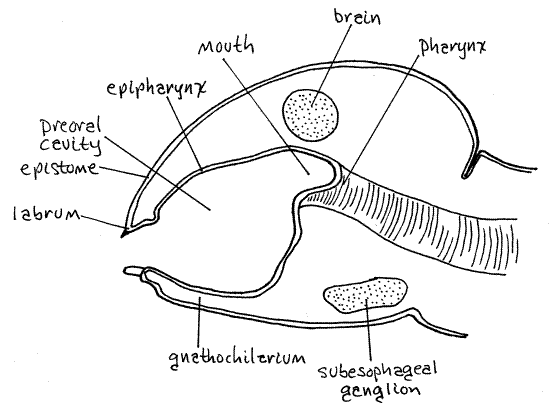
Preoral Cavity
With the gnathochilarium pushed aside look into the space thus revealed. It is the preoral cavity (Fig 7, 6), from which the mouth (which you will not see) opens. The millipede preoral cavity is bounded anteriorly by the labrum and epipharynx, laterally by the mandibles, and posteriorly-ventrally by the gnathochilarium. The epipharynx of the head capsule is the inner surface of the fold of cranial exoskeleton that forms the anterior part of the head. It extends from the labrum to the mouth and is mostly a thin unsclerotized membrane but it is not visible externally. The epistome is the outer surface of this fold.
Eyes
Although most millipedes have ocelli (Fig 20-11) laterally on the head, eyes are absent in polydesmids (Fig 3, 4).
Tömösváry Organ
A pair of sensory Tömösváry organs (TIM-ish-vary) is present on the head capsule just posterior and lateral to the antennae (Fig 4). They are small flat oval cuticular rings at the bases of the antennae. Tömösváry organs are thought to be hygroreceptors for assessing humidity, or possibly chemoreceptors.
Figure 7. Sagittal section of a millipede head to show the preoral cavity. (Redrawn from Snodgrass, 1951). Milliped32L.gif
Trunk
Segments, Diplosegments, and Rings
In polydesmids the trunk is divided into 20 trunk rings, of which a few are simple segments but most are double segments formed during development by fusion of two simple segments. Trunk rings composed of a single segment are called just that, segments, those consisting of two segments are known as diplosegments, and the term ring, or trunk ring, is used here in reference to all trunk annulations, segments and diplosegments alike. Segments have one pair of legs or no legs whereas diplosegments, as you would predict, have two pairs of legs. The presence of diplosegments is, of course, the reason for the name Diplopoda and it is an apomorphy of the taxon.
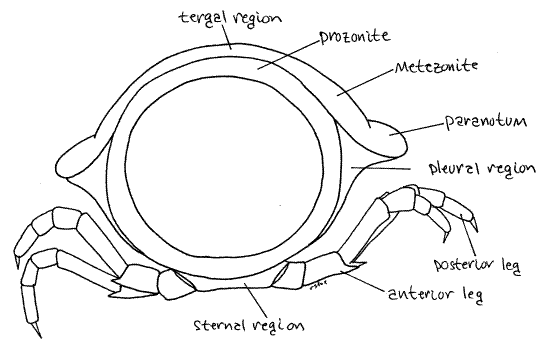
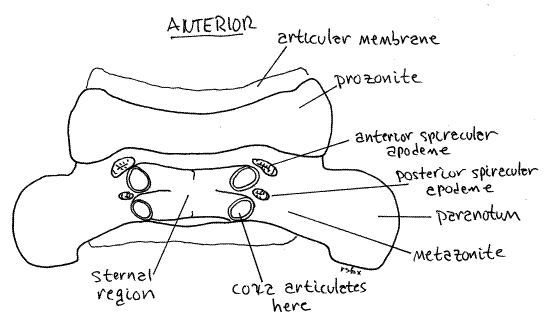
The exoskeleton of a typical arthropod segment consists of four sclerites (tergite, two pleurites, and sternite) joined to each other by flexible articular membranes to form a circle around the outside of the segment (Fig 16-1B). In millipedes the sclerites of each circle are fused together to form a rigid hoop, consisting of four regions with no flexible articular membranes or sutures separating them. The tergal region of the hoop is the largest and forms an arch covering the dorsum and upper sides of the ring (Fig 8). Each tergal region bears two lateral flanges, the paranota, that extend to the side (Fig 1, 8). The pleural and sternal regions are smaller. The sternal region is midventral and the coxae of the legs articulate with it. Each of the two pleural regions, which are intermediate in size, extends from a paranotum to the base of a leg (Fig 8).
Figure 8. Anterior view of a single diplosegment. Note that the body is circular, like that of a worm millipede, and the impression of flattening is due to the paranota. Milliped30L.gif
Ring 1 differs from the others in several respects. In the first place, it is a single segment that has no appendages. Further, its sclerites are not fused to form a rigid circle and instead are connected by flexible articular membranes. Finally, its tergite, tergite 1, is expanded to form a dorsal shield, the collum (= collar) (Fig 1, 20-9). Sternite 1 is small, yolklike, and straddles the ventral midline.
The first segment is usually considered to have no legs but it may look to you as if the first pair of legs belong to this first segment and, in fact, there is some discussion amongst diplopod specialists as to which segment the first pair of legs belongs.
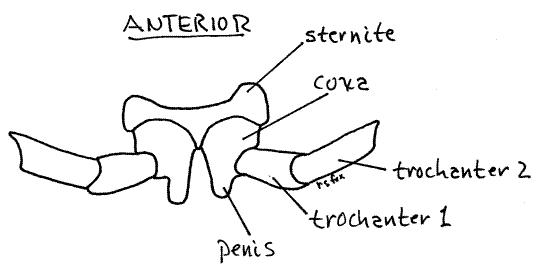
Rings 2-4 are also single segments but each has a pair of legs. Ring 2 bears the first pair of legs. The legs arise from sternite 2 (Fig 1) which is not rigidly fused to the other sclerites of the segment.
Rings 3-4 likewise are single segments and each has one pair of legs. The sclerites of these segments are fused to the other sclerites of their respective segments form rigid circles, one around each segment. These circles have no independent sclerites and instead are composed of tergal, pleural, and sternal regions (Fig 8, 20-10C).
Ring 3 bears the second pair of legs and is the genital ring. In both sexes it is the site of the gonopores. In females each of the two oviducts ends at a female gonopore in one of the two eversible pouchlike vulvae (sing. = vulva) just posterior and lateral to the coxa of the second pair of legs (Fig 2, 20-12 C, D). The vulva is surrounded by a sclerotized operculum.
In the male the genital ring bears a pair of penes (sing. = penis), each with a male gonopore through which a vas deferens from a testis opens. The penes (= genital papillae) are peg-like processes on the medial edge of the coxae (first article) of the second pair of legs (Fig 9). The second pair of legs is on ring 3.
In all myriapods except the chilopods (centipedes) the gonopores are situated anteriorly on the trunk, thus accounting for the name Progoneata (= front gonopore) for these taxa (millipedes, symphylans, and pauropods) which share this apomorphy. Chilopods, which have gonopores on a posterior segment are opisthogoneate (= back gonopore).
Figure 9. Ventral view of the second pair of legs of a male showing the penes. Milliped35L.gif
Ring 4 is another single segment with one pair of legs, the third. Ring 4 is the anteriormost ring to possess spiracles and, being a single segment, it has one pair of them. Diplosegments posterior to ring 4 have two pairs of spiracles (Fig 2, 11, 20-10B).
The spiracles are ventral and open from the lateral corners of oval spiracular apodemes on the anterolateral corners of the sterna supporting each pair of legs (Fig 11, 2). The coxae arise from these corners also and may hide the apodemes from your view. Move the leg aside, if necessary, to reveal the apodeme. The spiracle is the anterolateral corner of the apodeme and is difficult to discern. The spiracles of ring four are the largest of all and this is where you should look to see your first spiracles. Each spiracle opens into an internal atrium. A tuft of unbranched tracheae arises from each atrium. The tracheae extend to the tissues to deliver oxygen. Millipede atria are the hollow interiors of the apodemes. Apodemes in general are invaginations of the surface cuticle that function as an endoskeleton for muscle attachment (Fig 16-1E). These special spiracular apodemes have an additional function in respiration. With few exceptions millipedes are unable to close their spiracles making them subject to desiccation in dry environments.
The remaining trunk rings are diplosegments (= diplosomites). With the exception of rings 19 and 20, each has two pairs of legs articulating with the sternal region of a rigid cuticular hoop enclosing the double segment (Fig 8, 1, 2). Diplosegments are formed in the embryo by the fusion of two adjacent segments and accordingly have two pairs of legs, two pairs of ganglia, two pairs of ostia, two pairs of spiracles and, in the embryo only, two pairs of coelomic pouches.
The exoskeleton of each diplosegment is conspicuously divided into an anterior prozonite and posterior metazonite by a transverse suture (Fig 10, 11, 1). The paranota are on the metazonites and the two pairs of legs likewise arise from two sterna on each metazonite. Each metazonite bears two sternites and two pairs of legs. The terga of the metazonites are expanded laterally by the paranota so that the metazonites are much wider than prozonites. The prozonite of each ring telescopes into the metazonite of the preceding ring and is encircled by it. These two regions do not correspond to the two original segments that fused to form the diplosegment.
Figure 10. Dorsum of ring 10, disconnected from the adjoining rings to reveal the prozonite. Milliped34L.gif
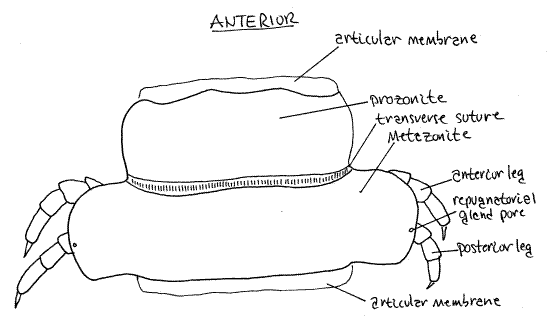
About half the paranota have a pair of small repugnatorial gland pores through which the repugnatorial glands (= stink glands) open to the exterior. These tiny, but clearly visible (with magnification), pores are very near the dorsal edges of the paranota of rings 5, 7, 9, 10, 12, 13, 15, 16, 17, 18, and 19) (Fig 1, 10, 20-10C). Focus on the margins of the paranota with high power (45X) of the dissecting microscope to find one. When you find a pore confirm that it is an opening by slipping a microneedle into it.
Ring 7 of males bears a pair of gonopods. These will be discussed later.
Ring 19, the preanal ring, is the last diplosegment. It has no legs and no spiracles but it does have repugnatorial gland pores.
The posteriormost unit of the trunk, which in polydesmids is ring 20 (= anal ring), is often called the telson (Fig 1, 14). It is not a true segment for it does not arise in the embryo from teloblast areas as do real segments. It has no legs but it bears the large anus, which will be discussed in more detail later, on its ventral surface.
In immature individuals of some millipedes there may be a zone of incomplete legless segments near the posterior end, between the last legged ring and the telson. This is the growth zone in which the mitotic teloblast areas are producing new segments.
Rings 1-4 are considered by some authors to constitute a separate tagma, the thorax. Justification for this is based on the single condition of the segments, legs that function as accessory mouthparts, absence of repugnatorial glands, absence of spiracles in all but one segment, and the presence of the gonopores. This view then combines all the diplosegments into another tagma, the abdomen.
Figure 11. Venter of ring 10 disconnected from the adjoining rings to reveal the prozonite. The legs have been removed for clarity. Milliped33L.gif
Leg
Legs are present on all trunk rings except 1, 19, and 20. Millipede legs are strongly homonomous, or similar to each other in size and morphology, with no regional specialization. The only exception being the extreme modification of the anterior pair of legs of the male 7 th ring, which become the intromittent organs known as gonopods, discussed below. Each leg (except the gonopod) is a walking leg used in locomotion. The legs extend laterally then ventrally to support the body as if in a hammock (Fig 8), rather like the legs of reptiles. Polydesmids have a total of 31 pairs of legs.
Each leg consists of seven articles. In the complete absence of consensus regarding the names, and even the number, of leg articles, the nomenclature of Snodgrass (1951) is followed here. Thus the articles are, in order from proximal to distal; coxa, trochanter 1, trochanter 2, femur, tibia, tarsus, and the clawlike pretarsus (Fig 12). Only the tips of the pretarsi contact the substratum when walking.
Gonopod
Male millipedes transfer spermatophores using one or two pairs of highly modified legs on ring 7 of the male (only). In polydesmids only the anterior pair of legs of the ring 7 is modified. These are intromittent organs, known as gonopods. Sperm transfer is indirect and employs the gonopods to move spermatophores from the male gonopores on his genital ring (3) to the female vulva on her genital ring. The complicated morphology of the gonopods (Fig 13, 20-12B) is species specific and serves as a mechanical species isolating mechanism. As is often the case in arthropods the shape of the gonopods is important in species level taxonomy and consequently is useful in species identification.
Figure 12. A typical leg, the anterior left leg of diplosegment 6. milliped24L.gif
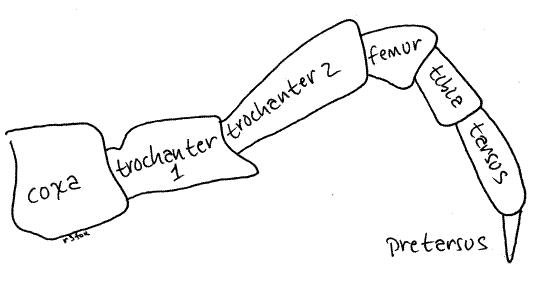
Figure 13. The gonopods of a male Pachydesmus. The rami of the telopodite are shaded to indicate their sclerotization. Milliped27L.gif
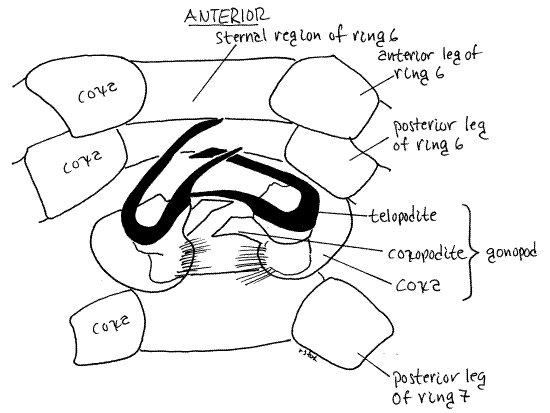
The gonopods are usually considered to be modified legs. Each gonopod consists of a bulbous basal coxa from which extends a small coxal endite and a large conspicuoustelopodite (Fig 13, 20-12B). (A telopodite is the movable part of an appendage.) In Pachydesmus the endite is weakly sclerotized and flexible. The large striking telopodite consists of two slender, gracefully curved, heavily sclerotized, rigid, brown rami. The posterior pair of legs on ring 7 are normal unmodified legs in polydesmids.
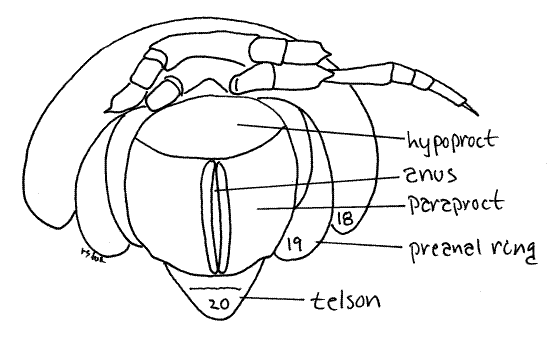
Telson
The posteriormost division of the body, following ring 19, is the triangular telson (= anal ring) (Fig 1). Not having arisen from embryonic teloblasts, is not considered to be a true segment. The large anus is on the venter of the telson protected on each side by a pair of clamshell-like sclerites, the paraprocts (Fig 14, = anal plates). A third sclerite, the hypoproct (= sub-anal scale) is immediately anterior to the anus on the venter. The two paraprocts swing open laterally to permit the extrusion of fecal pellets. With the paraprocts closed the anus is a long, midventral slit flanked by two cuticular ridges, which are the edges of the paraprocts. With your forceps open the paraprocts to expose the anus and reveal the spacious rectum, or hindgut, within. Fecal pellets are formed by the rectum prior to extrusion.
Figure 14. Ventral view of the posterior end of a female. Most of the legs of ring 18 have been omitted for clarity. Milliped26L.gif
References
Ax P. 2000. Multicellular animals, the phylogenetic system of the Metazoa. II. Springer Verlag, Berlin. 396pp.
Buck JB, Keister ML . 1950. Spirobolus marginatus, pp. 462-475 in Brown FA. Selected invertebrate types. Wiley, New York. 597pp.
Lawrence RF. 1984. The centipedes and millipedes of southern Africa, a guide. A.A. Balkema, Capetown, South Africa. 147 pp.
Hopkins SP, Read HJ. 1992. The biology of millipedes. Oxford Univ. Press, Oxford. 233 pp.
Kaestner A. 1968. Invertebrate Zoology, II. Arthropod relatives, Chelicerata, Myriapoda. Wiley Interscience, New York. 472pp.
Myers P, Espinosa R, Parr CS, Jones T, Hammond GS, Dewey TA . 2005. The Animal Diversity Web (online). University of Michigan Museum of Zoology. http://animaldiversity.org .
Shelley RM, Filka M. 1979. Occurrence of the milliped Pachydesmus crassicutis incursus Chamberlain in the Kings Mountain region of North Carolina and the Coastal Plain of South Carolina. Brimleyana 1:147-153.
Snodgrass RE . 1952 (reprint of 1971). A textbook of arthropod anatomy. Cornell University, Ithaca (Reprinted by Hafner Publ. Co, New York). 363pp.
Supplies
Dissecting microscope
Dissecting set with microdissecting tools
Flat millipede, living or preserved
Small dissecting pan (anchovy tin with wax bottom)
40% isopropanol