Invertebrate Anatomy OnLine
Limax maximus ©
Great Slug
with notes on Arion ater and Ariolimax columbianus
3jul2006
Copyright 2001 by
Richard Fox
Lander University
Preface
This is one of many exercises available from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises can be accessed by clicking on the links on the left. A glossary and chapters on supplies and laboratory techniques are also available. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Mollusca P, Eumollusca, Conchifera, Ganglioneura, Rhacopoda, Gastropoda C, Euthyneura, Pulmonata sC, Stylommatophora O, Sigmurethra sO, Aulacopoda iO, Limacacea SF, Limacidae F (Fig 12-125)
Mollusca P
Mollusca, the second largest metazoan taxon, consists of Aplacophora, Polyplacophora, Monoplacophora, Gastropoda, Cephalopoda, Bivalvia, and Scaphopoda. The typical mollusc has a calcareous shell, muscular foot, head with mouth and sense organs, and a visceral mass containing most of the gut, the heart, gonads, and kidney. Dorsally the body wall is the mantle and a fold of this body wall forms and encloses that all important molluscan chamber, the mantle cavity. The mantle cavity is filled with water or air and in it are located the gill(s), anus, nephridiopore(s) and gonopore(s). The coelom is reduced to small spaces including the pericardial cavity containing the heart and the gonocoel containing the gonad.
The well-developed hemal system consists of the heart and vessels leading to a spacious hemocoel in which most of the viscera are located. The kidneys are large metanephridia. The central nervous system is cephalized and tetraneurous. There is a tendency to concentrate ganglia in the circumenteric nerve ring from which arise four major longitudinal nerve cords.
Molluscs may be either gonochoric or hermaphroditic. Spiral cleavage produces a veliger larva in many taxa unless it is suppressed in favor of direct development or another larva. Molluscs arose in the sea and most remain there but molluscs have also colonized freshwater and terrestrial habitats.
Eumollusca
Eumollusca, the sister taxon of Aplacophora, includes all molluscs other than aplacophorans. The eumolluscan gut has digestive ceca which are lacking in aplacophorans, the gut is coiled, and a complex radular musculature is present.
Conchifera
Conchifera, the sister taxon of Polyplacophora, includes all Recent molluscs other than aplacophorans and chitons. The conchiferan shell consists of an outer proteinaceous periostracum underlain by calcareous layers and is a single piece (although in some it may appear to be divided into two valves). The mantle margins are divided into three folds.
Ganglioneura
Most Recent molluscs are ganglioneurans, only the small taxa Aplacophora, Polyplacophora, and Monoplacophora are excluded. Neuron cell bodies are localized in ganglia.
Rhacopoda
The mantle cavity is posterior in the ancestor although it may be secondarily moved to an anterior position by torsion. This taxon includes gastropods and cephalopods.
Gastropoda C
Gastropoda is the largest molluscan taxon and is the sister group of Cephalopoda. Gastropods are united by descent from a torted ancestor although many exhibit various degrees of detorsion. Many are coiled and asymmetrical but the ancestor was probably symmetrical. Gastropods are relatively unspecialized molluscs known collectively as snails. The univalve shell, present in the ancestral gastropod and in the majority of Recent species, is reduced or lost in many representatives. The flat creeping foot was inherited from their eumolluscan ancestors but gastropods have developed a distinct head with an abundance of sophisticated sense organs. The originally posterior mantle cavity has become anterior as a consequence of torsion, although detorsion has reversed this condition in many. Gastropods were originally gonochoric and most remain so but many derived taxa are hermaphroditic. Most are marine but many taxa have invaded freshwater and the only terrestrial molluscs are gastropods. Most have a single gill, atrium, and nephridium but the most primitive representatives have two of each. Only one gonad, the right, is present. The ancestor probably had an operculum. The nervous system is streptoneurous (twisted by torsion).
Euthyneura
Euthyneurans are derived prosobranchs thought to share a common mesogastropod ancestor. Euthyneura includes the no longer recognized taxa Opisthobranchia and Pulmonata. The asymmetrical streptoneurous nervous system created by the torsion that defines Gastropoda is wholly or partly reversed by detorsion. The result is a secondarily symmetrical, euthyneurous nervous system. Euthyneurans are hermaphroditic, exhibit a tendency to reduce or loose the shell, tend to be bilaterally symmetrical, and usually lack an operculum.
Pulmonata sC
The old subclass Pulmonata is no longer considered to be monophyletic and has been abandoned by zoologists as a formal taxon. Nevertheless it remains pedagogic convenience and is used here informally. Pulmonates are gastropods adapted for terrestrial and amphibious habitats. They are derived from prosobranch ancestors and have one atrium and one nephridium. The gill has been lost, however, and the mantle cavity is a heavily vascularized lung adapted for respiration in air. There is typically a coiled shell but some, the slugs, have reduced and internalized the shell and are not coiled and are secondarily symmetrical. Pulmonates are hermaphroditic and do not have an operculum. They are found in terrestrial habitats as well as near-shore, intertidal marine and freshwater environments.
Stylommatophora sC
Stylommatophora, the higher pulmonates, is the largest pulmonate taxon and includes the slugs and most terrestrial snails. (Aquatic pulmonates belong to Basommatophora.) Two pairs of tentacles are present and the eyes are at the tip of the posterior pair. Terrestrial snails are coiled, asymmetrical, and have a shell but slugs are uncoiled, symmetrical, and have lost or reduced the shell.
Laboratory Specimens
Terrestrial slugs belonging to several pulmonate families occur in moist areas throughout most of temperate North America. Many of the most widespread species, especially those common near human habitations, are introductions from Europe. Several genera have members achieving lengths in excess of 10 cm and are eminently suitable for use in teaching laboratories as examples of pulmonate gastropods.
Slugs, of course, are not typical gastropods, or even typical pulmonates, but they are nevertheless good subjects for an introduction to pulmonate anatomy. The extreme reduction of the shell and absence of coiling, although derived and atypical characteristics for a gastropod, greatly simplify the dissection and facilitates an understanding of the major features of pulmonate anatomy such as the lung and hermaphroditic reproductive system.
The organs are discrete, well defined, color-coded (in life), and easily recognized. Most are in the large accessible hemocoel that serves as the body cavity. That the dissection is far simpler than that of a coiled and shelled gastropod, such as Helix, and recommends it as an example of pulmonate anatomy, especially if studied in comparison with a more typical shelled and coiled prosobranch gastropod.
In North America, the species most likely to be used for this purpose are Limax maximus, Arion ater, and Ariolimax columbianus. Arion and Ariolimax belong to the same higher taxa (Arionacea SF, Arionidae F) and are similar to each other but less like Limax. Limax and Arion are European introductions but Ariolimax is a native species.
Limax maximus, the great slug, reaches 12 cm in length and is grayish with irregular black blotches and mottling. It is a common species in lawns, gardens, and moist basements or garages and it occurs in Europe and on the east and west sides of the North American continent (photo at: www.hiltonpond.org/ThisWeek000608.html ).
Arion ater, the black slug, is less widely distributed but may be very common in moist areas of the North American west coast. It reaches lengths of almost 15 cm and is dark brown, red, or black with a bumpy body (photo at: www.biopix.dk/Photo.asp?Photold=3774 ).
Ariolimax columbianus, the banana slug, is native to the west coast of North America where it is common in moist forests and may reach 15 cm in length. Its color is variable but is usually yellow, yellowish olive green, and often mottled (photo at: www.walnet.org/stanley_woods/slugz/bananaslug.html ).
Several native species in Philomycidae also exceed 10 cm and are usually restricted to relatively undisturbed habitats.
Limax and Ariolimax are available preserved from supply houses. Living slugs are not usually available commercially but in many parts of the North American continent, such as the west coast and the southeast, sufficient specimens to supply a small class can be collected locally. All three species are attracted to bowls of dry cat food (Purina Main Meal, or similar) left in dark moist areas in the summer or fall. They are nocturnal and inhabit moist places, becoming inactive during the winter.
Limax can be maintained in the laboratory in glass aquaria with a few centimeters of moist soil on the bottom. A few bricks or stones will provide a refuge for the nocturnal slugs. All three species can be maintained on the cat food mentioned above.
This exercise is written for Limax maximus with parenthetical comments pointing out features in which Arion and Ariolimax differ from Limax. Limax is marginally a better subject for this study because of the simpler topography of its gut. The fact that it is an introduced, rather than native species, also argues for its use in the laboratory in preference to the native Ariolimax. Either living or preserved specimens can be used. Living specimens are preferable to preserved.
Dissecting pans of convenient size can be made by pouring a wax bottom into old-fashioned aluminum ice trays or kippered herring cans.
External Anatomy
Observe the major external features of a living, unrelaxed specimen crawling in a large culture dish with about 1mm of water in the bottom. A preserved specimen can be used if necessary. The body is elongate, is not coiled, and is bilaterally symmetrical. The head is at the extreme anterior end and bears two pairs of tentacles and a pair of eyes. The long, oval foot is the ventral surface of the animal. The visceral mass lies dorsal to the foot and posterior to the head. The back of the visceral mass is the notum. A large opening, the pneumostome, on the right side of the anterior visceral mass opens into the lung.
Anesthetization
If you are using a living slug, it must be anesthetized prior to the dissection. This is necessary for humane reasons and to insure that the muscles are relaxed and uncontracted. Preserved slugs, of course, require no relaxation.
Living slugs, like most molluscs, are difficult to anesthetize in a relaxed condition with the muscles relaxed. The best method is to place the slug in a culture dish and gently cover it with carbonated water (club soda). Use a dish large enough for the animal to extend to its full length. Do not move the dish or disturb it in any way until the animal has succumbed to the anesthetic and can no longer respond to stimuli. The tentacles will probably be partially retracted and the head may be withdrawn, especially in Arion.
Once anesthetized, place the slug in a small wax-bottomed dissecting pan and cover it with carbonated water. Put the pan on the stage of a dissecting microscope and conduct the dissection under magnification.
Body
Study a relaxed, freshly killed, or preserved specimen. The body is elongate and thick, shaped like a short, fat worm or, not surprisingly, like a slug. Like the bodies of other molluscs it is composed of a head, foot, mantle, and visceral mass but there is no external shell.
The foot is the broad, flat, creeping surface occupying almost all the ventral side of the animal. The head is the anterior end of the animal and is relatively small and short (Fig 1, 12-28E). The visceral mass is most of the body. It sits dorsal to the foot and is almost everything behind the head. The upper surface, or back, of the visceral mass is called the notum. Themantle is the body wall of the notum. A conspicuous saddle-like area of body wall at the anterior end of the notum is actually a double fold of the mantle.
Foot
The foot occupies almost the entire ventral length of the body and all other body regions are above it. It is broad, muscular, and flat and used for creeping. Its ventral surface, or sole, bears two shallow, longitudinal grooves which separate a single median band from two lateral bands. The median band is ciliated. (The grooves are not present in Ariolimax or Arion.) The lateral edges of the foot are sharp and angular.
On each side of the animal, a little dorsal to the sole of the foot, a shallow, longitudinal pedal groove separates the foot from the rest of the body above it. These two pedal grooves connect anteriorly across the ventral midline with each other to form a transverse pedal groove which separates the head from the foot. A slime-producing pedal gland opens into this groove and secretes mucus to lubricate the path of the foot.
Figure 1. Dorsal view of a great slug, Limax maximus from Greenwood, South Carolina. Pulmon15La.gif
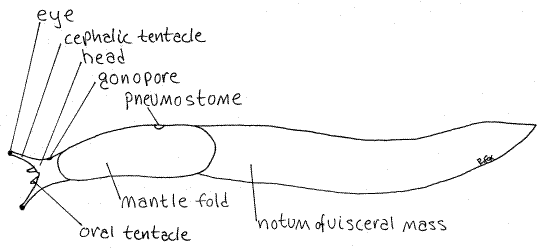
Head
The head is the anterior end of the body (Fig 1). It is separated from the foot by the transverse pedal groove described above. This groove extends a short distance dorsally on the sides of the body but does not encircle the animal. Dorsally there is no clear separation between the head and the visceral mass posterior to it. The head is connected to the visceral mass by an indistinct slightly narrower region called the neck. The head bears the median anteroventral mouth, just anterior to the transverse pedal groove.
The head is equipped with two pairs of sensory tentacles. The cephalic tentacles are dorsal and are larger than the ventral oral tentacles (Fig 1). The cephalic tentacles bear an eyeat the distal tip and are sometimes called ommatophores (ommat=eye, phore=to carry). The oral tentacles are mechano- and chemosensory.
Both pairs are completely retractile and, in fact, may be retracted in your specimen. If so, look at a healthy, unrelaxed specimen to find them. Tentacle retraction is accomplished by inversion. Each tentacle is a hollow diverticulum of the body wall, like the finger of a glove. The tentacle is pulled into the body by a long retractor muscle inserted on the inside surface of its distal end. Contraction of this muscle pulls the tip of the tentacle into the head and turns the tentacle outside in.
>1a. If a living, unrelaxed specimen is available, stimulate each of the tentacles and the head in turn by touching them gently with the tip of a probe or teasing needle and watch the response. Can these structures be retracted independently of each other or must all be withdrawn simultaneously? Does the degree of response seem to be related to the strength or frequency of the stimulus? Use a dissecting microscope or hand lens to observe the mechanism by which a tentacle is retracted. Can you see its tip being pulled into its core by its retractor muscle? <
Visceral Mass
The visceral mass is long and narrow and accounts for most of the body, except for the head, above the foot (Fig 1). It is thick and wide and shaped in cross section like a loaf of bread. Posteriorly it bears a distinct, longitudinal, median, dorsal, ridge, or keel. (There is no distinct keel in Arion). Its surface bears closely space slime grooves. (The pattern of slime grooves and ridges is especially evident in Arion.)
The slightly narrower region of the visceral mass immediately posterior to the head is the neck. (In Arion there is a large, mucus-secreting caudal gland at the posterior tip of the visceral mass.)
Mantle
The mantle is the dorsal body wall of the visceral mass. Immediately posterior to the neck it is folded to form a low, flat, oval mass of tissue that, as in other molluscs, encloses the mantle cavity and is its roof.
Anteriorly, and to a lesser extend laterally, the roof of the mantle cavity is free and unattached to the underlying visceral mass. In these areas it forms a large free lobe lying on the surface of the anterior visceral mass. Posteriorly the roof is attached directly to the back, or notum, and does not form a lobe. Lift the edges of the roof anteriorly, laterally and posteriorly to see this. (There is a small posterior lobe in Ariolimax and Arion.) The head can be retracted beneath the anterior lobe.
Mantle Cavity
The mantle cavity of pulmonates is a lung, or pulmonary sac, and is almost completely enclosed. It does not have the wide anterior opening typical of prosobranch snails. The mantle skirt grows to the notum and neck leaving only a small pore, the pneumostome, connecting the lung with the exterior. This opening is located near the edge of the mantle on the right side. InLimax and Ariolimax it is on the edge of the posterior half of the mantle but in Arion it is a little anterior to the middle of the mantle. The pneumostome is equipped with muscles and can be opened and closed. The mantle cavity does not contain a gill, hypobranchial gland, or osphradium as it does in prosobranchs but its roof is heavily vascularized and functions as a lung for aerial respiration.
The anus is located on the right side immediately inside the pneumostome. It will be easier to see after you open the lung. It is connected to the lateral border of the mantle and to the dorsal edge of the pneumostome by a deep, oblique anal groove extending ventrally from the edge of the pneumostome. The groove runs from the pneumostome and anus anteroventrally. Use magnification and a probe to explore the proximal end of the groove and find the anus. The nephridiopore is also located in this area but you probably will not see it.
The common gonopore serving both male and female reproductive systems is located on the right side of the head posterior to the bases of the two right tentacles. (It is directly ventral to the pneumostome in Arion.) The gonopore is relatively large but it is inconspicuous nevertheless and magnification will probably be required to find it.
" Orient the slug in a long narrow dissecting pan with its ventral surface down. Use four #1 insect pins inserted at 45 ° angles through the sides of the foot (not through the visceral mass) to anchor it in place. The slug should be immersed in carbonated water, if alive, or tapwater, if preserved. The dissection should be conducted with magnification.
Use fine scissors to open the lung. Start at the ventral margin of the pneumostome and cut anteriorly along the line separating the mantle cavity roof from the visceral mass. Lift the roof out of the way and cut as close to the visceral mass as possible but do not cut through the dorsal body wall (which is the floor of the lung). Cut around the anterior roof and posteriorly on the left side. Do not cut across the posterior border of the roof, leaving it as a hinge.
Return to the pneumostome and cut posteriorly from its lower margin, still cutting as close to the visceral mass as possible. Again, do not cut transversely across the posterior mantle.
When you have finished, the roof will remain attached posteriorly and can be deflected in that direction to reveal the lung.
Lift, deflect, and pin back the roof and look at the cavity it concealed. This space is the mantle cavity, or lung. In it can be seen the kidney and heart. The floor of the mantle cavity is the dorsal body wall. The roof is the double fold of mantle. The organs of the lung are all in its roof. The pericardial cavity and heart are on the left with the atrium anterior to the ventricle. The kidney and rectum are on the right. Most of the inner surface of the roof is heavily and conspicuously vascularized for gas exchange.
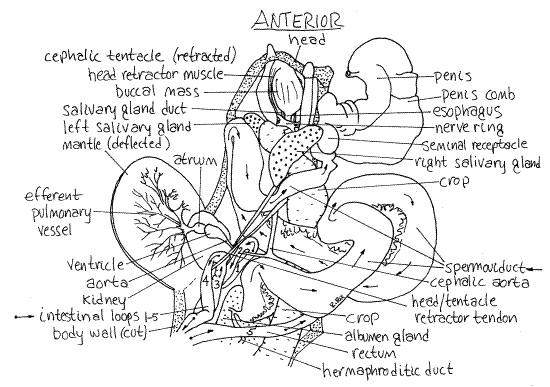
Figure 2. Dorsal view of a dissected Limax maximus. The limbs of the intestinal loops are numbered. Pulmon16La.gif
The large, creamy white (in living Limax, but pale brownish gray in Ariolimax) nephridium, or kidney, is the most conspicuous structure in the lung (Fig 2). It bulges into the posterior cavity and occupies most of the space there. Look for the inconspicuous ureter which exits the posterior right end of the nephridium and extends to the nephridiopore beside the anus. (In Arionthe nephridium does not bulge strongly into the mantle cavity. It is pale brown and lies on the roof of the cavity. There is a narrow yellowish gland beside the left side of the nephridium of Arion.)
If you did not see the anus earlier, look for it now on the inside of the dorsal lip of the pneumostome. It is a large slit. You can also see the rectum running a short distance across the roof of the posterior right lung to enter the visceral mass. The rectum crosses the roof of the cavity obliquely to reach the anus. Find the anus at the end of the rectum. Insert a scissors in the anus and open a few millimeters of the rectum to demonstrate its tubular nature. Its inner walls are longitudinally ridged. The ureter runs beside the rectum and opens via the nephridiopore near the anus.
The thin, membranous pericardium is attached to the ventral surface of the left side of the nephridium and encloses the pericardial cavity. (In Ariolimax the nephridium almost entirely covers and obscures the pericardium.) With the roof of the lung deflected, the pericardium lies on top of the nephridium. (In Ariolimax the pericardium can be seen beside and under the left side of the nephridium.) The pericardium bulges into the mantle cavity also and is translucent and white. (It does not bulge in Ariolimax and is not an obvious feature of the mantle cavity. InArion the pericardium is embedded in the nephridium and is obscure.) The large ventricle and smaller atrium are contained within the pericardial cavity but their shapes are discerned only with difficulty through its nearly opaque walls. (They cannot be distinguished in Ariolimax.)
" With fine scissors, carefully open and remove the pericardium to expose the heart but do not damage the vessels entering or leaving it.
With the pericardium out of the way it is easy to distinguish the atrium from the ventricle. In its normal position, with the mantle roof in place, the atrium is anterior to the ventricle (Fig 2). The atrium is smaller than the ventricle.
The efferent pulmonary vessel, which drains the pulmonary plexus of vessels in the roof of the lung, enters the atrium anteriorly whereas the aorta exits the ventricle at the opposite (posterior) end of the heart (Fig 3). (There is no single afferent pulmonary vessel in Ariolimax but the aorta is large and conspicuous.) This end of the ventricle lies against the floor of the mantle cavity and the aorta penetrates this floor and quickly divides into an anterior cephalic aorta (Fig 3), which you may be able to see through the transparent floor and a posterior visceral aorta, which you cannot see yet.
The lung of pulmonates is the respiratory organ and its roof and extreme right floor are heavily vascularized. (The vascularization is not so heavy or obvious in Ariolimax. It is the floor of the lung that is heavily vascularized in Arion.) In Limax the pulmonary blood vessels in the roof and right floor are easily seen and form an intricate network of afferent and efferent pulmonary vessels draining into the atrium of the heart. This network is the pulmonary plexus. The afferent vessels bring unoxygenated blood from the hemocoel to the lung whereas the efferent vessels conduct oxygenated blood from the lung to the atrium.
Most of the floor of the lung is not vascularized. It is part of the dorsal body wall, or notum, and is tough and transparent.
" Open the posterior end of the lung roof with a shallow dorsal incision and find the small, oval shell.
The shell is embedded in the posterior lung roof which entirely conceals it. Remove the shell, place it in a small dish of water, and observe it with the dissecting microscope. It is composed of protein and calcium carbonate and bears asymmetric growth lines.
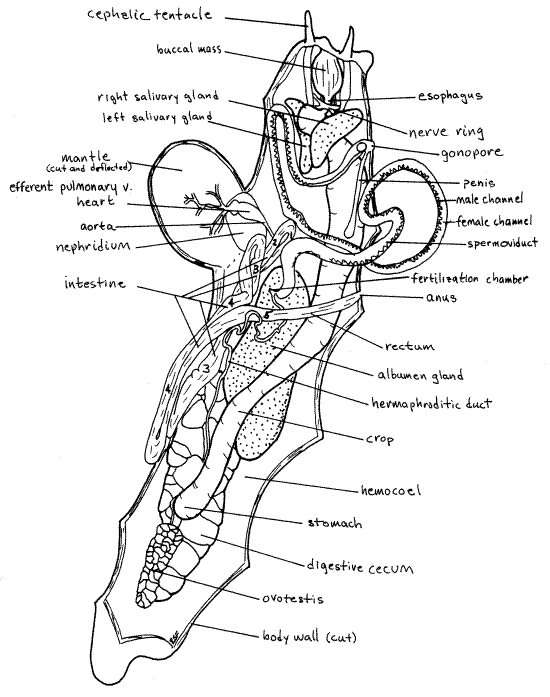
Figure 3. Dorsal view of the head and mantle cavity of a dissected Limax maximus. The penis is everted in this specimen. Unadorned arrows indicate the direction of gametes through the gonoduct. Tailed arrows show the movement of food through the loops of the intestine. The limbs of the intestinal loops are numbered. Pulmon17La.gif
(In Ariolimax the shell is a thin, flexible, oval, transparent plate in the lung roof dorsal to the nephridium. Free the nephridium from the mantle by carefully cutting the thin connective tissue holding it in place. Note that there are two regions of the kidney. One you have already seen bulging into the mantle cavity whereas the other adheres tightly to the mantle. Free this tightly adhering region and look for the shell dorsal to it.)
Internal Anatomy
" Free the kidney from the floor of the lung as necessary and deflect the roof to the left. Leave the ventricle attached to the aorta. Do not remove the mantle or cut along its posterior margin.
Make a median, dorsal, longitudinal incision through the body wall (notum) from a position between the cephalic tentacles posteriorly to the posterior tip of the visceral mass. It is best to make this incision under magnification to insure that you do not cut deeper than the body wall. Pin the body wall aside with # 1 insect pins as you proceed. Be careful of the internal organs as some of them are delicate and close to the surface. The rectum especially is easily destroyed. It is a diaphanous, delicate, flat tube extending obliquely across the body cavity from left to right. It looks like a narrow mesentery and might inadvertently be removed.
Preview
The cavity thus exposed is the hemocoel (Fig 2). It is not a coelom but is large and spacious and extends for the entire length of the animal. Locate some of the major organs in the hemocoel for later use as landmarks. The muscular, ovoid mass filling most of the space in center of the head is the buccal mass (Fig 2). Inside it are the pharynx, radula, and jaw, which you will see later.
On either side of the buccal mass are two dark, longitudinal bands. These are the cephalic tentacles and their retractor muscles.
The enormous, lobulated, pale cream (in life) digestive cecum occupies most of the posterior space in the hemocoel. (The digestive cecum of Ariolimax is brown.)
The smaller, dark but also lobed ovotestis sits dorsal to the digestive cecum at the extreme posterior end of the hemocoel (Fig 2). (The ovotestis of Ariolimax is a small yellowish brown, more or less spherical organ near the middle of the body.)
A large pale yellow albumen gland (white in Ariolimax) dominates the middle region of the hemocoel (Fig 2, 3). It is ventral to most of the other structures in this region. Between the anterior end of the albumen gland and the buccal mass the hemocoel is nearly filled with a very thick, white, spermoviduct, or common male and female gonoduct (Fig 2, 3). It is very thick and appears to be a many-lobed gland rather than a duct. It is, in fact, a double tube connecting the ovotestis and albumen gland with the gonopore on the right side of the head. The large whitepenis can be seen extending into the interior from the common gonopore (Fig 3).
The long, wide, thin-walled, tubular crop runs posteriorly from the buccal mass on the right ventral side of the hemocoel (Fig 2, 3). It ends near the posterior end of the digestive cecum. Numerous transparent arteries can be seen running through the hemocoel. (The arteries of Ariolimax are opaque white, abundant, and highly branched.)
Limax Reproductive System
Because of its great size, it is best to study the reproductive system first and then remove it to make it easier to see the remaining organs. Fertilization is internal and the reproductive system is complex, as it inevitably is in hermaphrodites with internal fertilization. Many parts are shared between the male and female systems. (The reproductive systems of Ariolimax andArion differ from that of Limax and will be described separately.)
The ovotestis is a hermaphroditic gonad. It is a dark, brownish, lobed organ at the extreme posterior end of the hemocoel (Fig 2). It occupies a gonocoel, which is remnant of the coelom. It is located on the dorsal side of the posterior end of the digestive cecum.
A small, but distinct and easily found, hermaphroditic duct emerges from the anterior end of the ovotestis (Fig 2). It is a combined male and female gonoduct. Trace it as it runs anteriorly over the stomach and crop and under the dorsalmost lobe of the digestive cecum. Do not confuse it with the similar but transparent and branched visceral aorta beside it. The hermaphroditic duct is not branched.
The hermaphroditic duct runs across the dorsal surface of the large albumen gland from whose anterior end it receives a duct (Fig 2). The small fertilization chamber at the junction of the albumen duct with the hermaphroditic duct thought to be the site of fertilization.
The hermaphroditic duct continues anterior to the albumen gland as the larger spermoviduct (Fig 2, 3) characterized by greater separation of male and female channels. This anterior region of the gonoduct is much greater in diameter than the hermaphroditic duct. Most of the space in the middle and anterior hemocoel may be filled with the coils of this duct. The spermoviduct is incompletely separated into side by side male and female channels. The entire length of the duct is divided into a male channel, which contains spermatozoa and is intensely white, and a much wider, glandular female channel (Fig 2). The two channels are closely associated over most of the length of the duct.
As the spermoviduct approaches the gonopore on the right side of the head, it diminishes in diameter and the male and female channels separate completely from each other temporarily before rejoining to share the gonopore. A white, ovoid seminal receptacle is attached to the duct near the point where the female duct rejoins the male duct (Fig 3).
The distal end of male duct is wide and eversible and is the muscular, white penis (Fig 2, 3). This part of the duct can be turned inside out through the gonopore during mating and sometimes during anesthetization. When everted the penis is a thick tube with a wide, thin wing, the penis comb, along one border (Fig 3). The penis of your specimen is probably in its retracted resting condition inside the hemocoel. It is a large white tube (Fig 2).
The penis retractor muscle originates on the dorsal body wall near the posterior lung and extends to the penis where it inserts. Its tendon is easily traced across the hemocoel.
The glandular walls of the spermoviduct may be swollen and enlarged to such an extent that it is difficult to study other organs in the anterior hemocoel. You may remove the spermoviduct and discard it now if you wish. Be careful that you do not damage any other tissues, especially the transparent rectum, as you extract it. The albumen gland, ovotestis and proximal hermaphroditic duct should be left intact.
Courtship and Mating in Limax
Pulmonates are simultaneous hermaphrodites and, while cross fertilization is the rule, Limax sometimes self fertilizes. There is some inconclusive evidence of protandry in Limax. Courtship and mating of Limax maximus is elaborate, unusual, and well studied (Chase, 1972). During warm months pairs of sexually receptive individuals climb to some elevated position in a bush or tree and follow one another in a circle while licking each other for as much as 2 ½ hours. Eventually they entwine and drop from the perch and hang suspended, like amorous bungee jumpers, by a tough mucus strand that may be as much as 25 cm long (Fig 12-58A).
While entwined, each partner everts its penis to its full length, which may be 10 cm. The distal end of each penis wraps around the other penis and together they form a ball covered by the extended, winglike penis combs (Fig 12-58A). A mutual exchange of sperm occurs and the penes unwind and are retracted.
The slugs climb back up the mucus strand, often eating it as they climb. The sperm are stored initially in the seminal receptacle but eventually make their way upstream to the fertilization chamber where fertilization probably occurs. Eggs move downstream from the ovotestis and are fertilized in this chamber. Once fertilized the egg moves down the female channel of the spermoviduct, receiving numerous coatings from the albumen gland and secretions from the walls of the spermoviduct.
The eggs, when released, are transparent, colorless spheres about 4 mm in diameter. They gradually become cloudy and translucent and resemble pearls or spheres of the time-release fertilizer, Osmocote. The eggs are laid in the fall in clusters of 50-130 on moist ground under stones or other objects. Development is direct with tiny slugs emerging from the eggs. At least two years are required to achieve sexual maturity.
Ariolimax Reproductive System
The reproductive system is hermaphrodite, of course, and the common gonopore is located on the right side of the head. The system begins with a compact, globular, slightly yellowish to white, hermaphroditic gonad, the ovotestis, located near the middle of the hemocoel (Fig 4). It is dorsal and near the midline at about the level of the anterior end of the anteriormost lobe of the digestive cecum and the posterior margin of the lung roof.
A thick, but short, convoluted hermaphroditic duct exits the ovotestis and runs medially to the massive, highly lobulated albumen gland. This gland is white and occupies much of the space in the middle of the right side of the hemocoel. It obscures other structures in the area. It is largest in reproductively active individuals. The ovotestis lies to the left of it.
A very large, glandular spermoviduct exits the anterior end of the albumen gland and superficially appears to be simply a continuation of the gland. On closer inspection, however, it can be seen to be a thick convoluted tube. Anteriorly it looses its glandular character, decreases in diameter, and its tubular nature becomes more apparent. The spermoviduct consists of a combined sperm duct and oviduct, the lumina of which are only partially separated. The bright white sperm duct is easily distinguished from the larger, more glandular oviduct on whose surface it lies.
Distally the spermoviduct splits into separate sperm duct and oviduct. The sperm duct is small in diameter whereas the oviduct is large (Fig 4). At about this level a large, thin-walled, diaphanous sac, the copulatory bursa (= spermatheca), is attached to the oviduct by a sheet of connective tissue.
Follow the oviduct from the bifurcation of the spermoviduct. It continues anteriorly, soon receives the duct from the copulatory bursa, and then widens to form the short vagina, which extends to the common gonopore.
Return to the point at which the sperm duct diverges from the oviduct and follow the sperm duct anteriorly. Do not confuse the sperm duct with arteries in the area, which may be about the same size. Arteries branch, the sperm duct does not. The sperm duct runs anteriorly beside the oviduct and vagina, then turns medially and passes ventral to the vagina. Upon emerging on the left side of the vagina, the duct widens and extends in a great loop to the left side of the body. Here it enters the much larger and wider penis.
Figure 4. A dorsal view of the reproductive system of a banana slug, Ariolimax columbianus, from Charleston, Oregon. Pulmon18La.gif
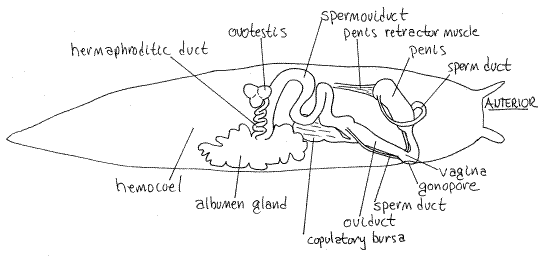
The penis is attached to the floor of the mantle cavity by a wide, flat penis retractor muscle. The penis itself extends obliquely across the anterior hemocoel to join the vagina just inside the gonopore. (You may wish to remove the albumen gland after completing the study of the reproductive system.)
Arion Reproductive System
The white ovotestis, enclosed in a transparent black sheet of connective tissue, is at the extreme posterior end of the hemocoel and is more or less spherical (Fig 5). It is surrounded by digestive cecum.
The conspicuous, white, convoluted hermaphroditic duct coils its way anteriorly from the ovotestis to join the white albumen gland near the middle of the body. The hermaphroditic duct is surrounded by coils of the intestine, digestive cecum, and crop.
From the albumen gland arises the very thick, white, convoluted, and glandular spermoviduct. When fully developed, this duct fills much of the space in the anterior hemocoel. It is much thicker than the hermaphroditic duct.
The spermoviduct makes its way anteriorly before dividing into a narrow sperm duct and a larger oviduct.
The sperm duct runs to the muscular penis from which arises a large, thin-walled sac, the copulatory bursa (= spermatheca). The oviduct runs anteriorly to the large, muscular vagina, which is joined distally to the penis. The penis and vagina share a common gonopore on the right side of the head.
Figure 5. Dorsal view of the reproductive system of Arion ater based on specimens from Charleston, Oregon. Pulmon19La.gif
Digestive System
Most slugs, including Limax, Ariolimax, and Arion, are herbivorous and feed on a variety of vegetation. Some, especially the exotic Limax and Arion, but not the native Ariolimax, can be voracious garden pests.
" Relocate the mouth at the anterior end of the body and insert one blade of a fine scissors into it and cut posteriorly through the dorsal wall of the buccal mass (Fig 2). This will open the mass with a median longitudinal incision. Be careful as you cut posteriorly into the buccal mass that you do not destroy the brain (Fig 2, 3). It is a nerve ring around the posterior buccal mass and anterior esophagus.
From your present viewpoint the nerve ring will appear as a bilobed white mass located on the posterior dorsal surface of the buccal mass. Avoid damaging it.
Inside the buccal mass you will find the lumen of the pharynx, into which the mouth opens. A large chitinous jaw with lateral teeth on each side arches over the lumen of the pharynx and may protrude from the mouth. The flexible, transparent, rasplike radula is located on the floor of the pharynx. Several head retractor muscles arise on the sides of the buccal mass and extend posteriorly through the hemocoel (Fig 3). They will be traced later.
>1b. Remove a piece of the radula, make a wetmount, and examine it with the compound microscope to see the abundant teeth and their arrangement. <
A narrow, short esophagus emerges from the posterior end of the dorsal buccal mass (Fig 2, 3). The two salivary gland ducts enter the gut laterally at the junction between the esophagus and pharynx (Fig 3). Trace the ducts posteriorly to the two large, white salivary glands (Fig 2, 3). (The salivary glands of Ariolimax may be yellow.) Do not damage the nerve ring through which these ducts pass. The shape and position of the glands is asymmetrical. The right gland is dorsal in the anterior hemocoel whereas the left gland is ventral. The right gland is easily seen from your present viewpoint but the left is obscured by the other organs in the region. (The asymmetry is even more pronounced in Ariolimax.) The right salivary gland is very large. It lies mostly on the dorsal left but curves around to the right also. Its large duct empties into the pharynx on the right, thus revealing that it is the right gland. The left gland is long and narrow and lies on the floor of the hemocoel. Its duct enters the pharynx on the left, of course. Do not confuse the salivary ducts with blood vessels in the area.
The esophagus passes posteriorly through the white circumesophageal nerve ring, or brain (Fig 2, 3). (In Ariolimax the nerve ring encircles the posterior pharynx.) The nerve ring is composed of two dorsal cerebral ganglia plus paired pedal, visceral, and pleural ganglia located lateral and ventral to the gut. Numerous large nerves radiate from the ganglia. Do not damage the nerve ring and its nerves.
The esophagus, salivary ducts, cephalic aorta, retractor muscles from the ventral tentacles and head all pass through the nerve ring. The head retractor muscles insert on the sides of the buccal mass and pass posteriorly through the ring, as you already know. The retractor muscles of the oral tentacles do likewise. The retractors of the cephalic tentacles do not pass through the ring.
Posterior to the nerve ring the oral tentacle retractors, cephalic tentacle retractors, and head retractors unite to form a single muscle and tendon that runs posteriorly to its origin on the body wall under the lung (Fig 3). The retractors are derived from the columellar muscle of the shelled ancestor. Notice that the penis retractor is not a part of this system and originates independently but nearby.
Posterior to the nerve ring the esophagus empties into the large, long, translucent, and very thin-walled crop (Fig 2,3). (In Ariolimax and Arion the intestine and digestive cecum spiral around the crop and obscure your view of it.) The contents of the crop are seen clearly through its walls, making it the color of the slug's most recent meal.
The crop extends posteriorly to the anterior end of the ovotestis where it expands slightly to become the short stomach (Fig 2). (In Ariolimax and Arion the stomach is well posterior to the ovotestis, in about the middle of the digestive ceca. It is a blind thick-walled pouch with crop and intestine attached to its anterior end. The large ducts from the digestive gland empty into it.)
The posterior crop and stomach are surrounded by the two large, lobed digestive ceca (Fig 2). Large ciliated ducts from the ceca communicate with the stomach. Intracellular digestion occurs in the ceca.
>1c. Open the stomach and wash away its contents to demonstrate the connections with the digestive ceca. <
The intestine exits the left side of the stomach. It is very long and makes 2½ longitudinal loops consisting of five limbs, before it empties at the anus. (In Ariolimax and Arion the intestine spirals around the crop in transversely, rather than in longitudinally oriented loops.) Limb 1 begins at the stomach and runs anteriorly on the ventral left side of the digestive ceca. It cannot be seen in dorsal view without turning the digestive ceca over. (Its connection with the stomach is not visible in Figures 2 and 3.) Limb 1 has relatively thick, opaque walls and usually contains food and is round in cross section. It extends anteriorly to the level of the kidney in the posterior mantle cavity where it loops around the cephalic aorta and reverses direction, becomes limb 2, and heads posteriorly again (Fig 2, 3).
Limb 2 parallels limb 1 and is still on the left but in a more dorsal position where it is a little easier to see. Its walls are slightly more transparent than those of limb 1 and fecal pellets can be seen in it if the slug has fed recently. It runs posteriorly along the dorsal surface of the anterior digestive cecum and then reverses direction and courses anteriorly again, now as limb 3 (Fig 2, 3).
Limb 3 runs anteriorly until it reaches the level of the lung where it loops around the tendon of the combined head and tentacle retractors and starts posteriorly as limb 4 (Fig 2, 3).
Limb 4 extends posteriorly on the left side of the hemocoel and digestive ceca. It tends to be flattened and becomes very thin-walled and diaphanous before it completes its posterior journey. It lies on the dorsal side of the digestive cecum. At about the level of the stomach the intestine reverses direction for the last time and becomes limb 5, which is the rectum.
The rectum is also thin-walled, transparent, and delicate. It is easy to overlook it or mistake it for a mesentery (of which there are none in a hemocoel) and you may have destroyed it already. (In Ariolimax and Arion the intestinal walls are more robust than in Limax.)
The rectum adheres to the posterior end of limb 4 for the first part of its anterior course and then breaks free and heads diagonally across the dorsal surface of the digestive cecum and albumen gland to end at the anus near the pneumostome (Fig 2). It begins on the left and crosses the hemocoel to the anus on the right.
Hemal System
The heart consists of an atrium and a ventricle located in the lung on its roof. In the undissected animal the atrium is anterior and the ventricle posterior (Fig 3). The efferent pulmonary vessels in the roof of the mantle drain into the atrium. The aorta exits the ventricle to deliver oxygenated blood to the hemocoel.
" If the pericardium has not been opened, open it now but do not damage the vessels attached to the heart.
Relocate the atrium and find the entrance of the efferent pulmonary vessel into its anterior end. The larger ventricle at the posterior end of the pericardial cavity pumps oxygenated blood into the aorta emerging from its narrow posterior end (Fig 3). The aorta penetrates the floor of the lung and enters the hemocoel. Follow it as it does so, or find it inside the hemocoel. It soon branches into two large vessels, one supplying the anterior body with oxygenated blood, the other supplying the posterior body.
Of these, the cephalic aorta, with its many branches, runs anteriorly and passes through the nerve ring (Fig 3). The visceral aorta runs posteriorly beside limb 2 of the intestine. You may have seen it earlier beside the hermaphroditic duct. It also has many branches. The arteries terminate in fine vessels which open into the venous hemocoel system. Numerous venous channels return blood from the hemocoel to a circular afferent pulmonary vessel in the roof of the lung. This vessel runs around the periphery of the lung. Smaller afferent vessels from the circle supply the capillaries of the pulmonary plexus. Abundant small efferent vessels drain these capillaries to the one large efferent pulmonary vessel which enters the apex of the atrium (Fig 3). The vessels are easy to see although some may have been damaged by your earlier activities. You may want to trace some of the branches of the two aortae.
Excretory System
The nephridium, or kidney, is located on the roof and posterior wall of the lung to the right of the heart (Fig 2, 3). (In Arion it is dorsal to the heart.) It is a metanephridium connected with the pericardial cavity (a coelomic space) by the renopericardial duct and with the exterior by the ureter. Most of the nephridium is a sac with thick, folded walls.
The ureter of Limax, as in all Sigmurethra sO, is sharply curved on itself. It exits the kidney and runs posteriorly along its medial edge then bends sharply and runs anteriorly, paralleling and touching the rectum. The nephridium was seen earlier in the posterior mantle cavity. The ureter and nephridiopore are difficult to find.
Nervous and Sensory Systems
The nervous system of Limax is easily studied. It is cephalized and euthyneurous, having reversed the effects of torsion, and is almost symmetrical.
The circumesophageal nerve ring, or brain, encircles the esophagus (Fig 3). All the major ganglia (cerebral, pleural, pedal, parietal, and even visceral) are present in the nerve ring.
The cerebral ganglia are dorsal to the gut whereas the remaining ganglia are ventral. On each side two connectives join the cerebral ganglion with the ventral ganglia of that side. Adjacent ganglia of the ring are fused together and lack visible connectives.
A pair of small buccal ganglia is present on the posterior wall of the buccal mass. Each is connected to the cerebral ganglia via a connective and to the other buccal ganglion by the buccal commissure.
Many nerves arise from the ganglia of the nerve ring. An easily recognized optic nerve runs from the cerebral ganglion to the eye at the end of each cephalic tentacle. If the tentacle is retracted, the eye will not be at the end. You can rectify this by slipping your finest forceps into the retracted tentacle and pulling it “right side out”.
Find the cephalic and oral tentacles on the anterior surface of the head and look inside the wall of the head for the dark tentacle retractor muscle associated with each tentacle. Posteriorly these muscles share a common origin with the head retractor muscles near the floor of the lung. These muscles are all derived from the right columellar muscle of the shelled mesogastropod ancestors of slugs.
References
Bullough WS. 1958. Practical Invertebrate Anatomy. MacMillan, New York. 483p.
Carmichael EB. 1937. Culture methods for Limax flavus in Needham JG. (ed), Culture Methods for Invertebrate Animals. Comstock, Ithaca. 590p.
Chase LM. 1952. The aerial mating of the great slug. Discovery 13 (11):
Hyman LH. 1967. The Invertebrates: Mollusca I. McGraw-Hill, New York. 792p.
Kofoid CA. 1895. On the early development of Limax. Bull. Mus. Comp. Zool. Harvard 27:1-119, pls. 1-8.
Petrunkevitch A . 1916. Morphology of Invertebrate Types. MacMillan, New York. 263p.
Pierce SL, Maugel TK. 1987. Illustrated Invertebrate Anatomy. Oxford, New York. 307p.
Simpson GB. 1901. Anatomy and physiology of Polygyra albrolabris and Limax maximus and embryology of Limax maximus. Bull. New York State Mus. 40(8):236-314.
Ruppert EE, Fox RS, Barnes RB. 2004. Invertebrate Zoology, A functional evolutionary approach, 7 th ed. Brooks Cole Thomson, Belmont CA. 963 pp.
Supplies
Dissecting microscope
Compound microscope
Slides and coverslips
Dissecting pan
# 1 stainless steel insect pins
Living or preserved large slug
Carbonated water (club soda)
Dissecting set with microdissecting tools
Preserved Limax is available from Wards and Carolina
Preserved Ariolimax is available from Carolina (as " Limax or Ariolimax")