Invertebrate Anatomy OnLine
Leiobunum ©
Harvestman
19jun2006
Copyright 2004 by
Richard Fox
Lander University
Preface
This is an exercise from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises can be accessed by clicking on the links to the left. A glossary and chapters on supplies and laboratory techniques are also available. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Arthropoda P, Chelicerata sP, Euchelicerata, Arachnida C, Lipoctena, Apulmonata, Holotracheata, Cryptoperculata, Opiliones O, Phalangida, Eupnoi, Phalangioidea SF, Phalangiidae F(Fig 16-15, 18-44, 18-47)
Arthropoda P
Arthropoda, by far the largest and most diverse animal taxon, includes chelicerates, insects, myriapods, and crustaceans as well as many extinct taxa such as Trilobitomorpha. The segmented body primitively bears a pair of jointed appendages on each segment. The epidermis secretes a complex cuticular exoskeleton which must be molted to permit increase in size. Extant arthropods exhibit regional specialization in the structure and function of segments and appendages but the ancestor probably had similar appendages on all segments. The body is typically divided into a head and trunk, of which the trunk is often further divided into thorax and abdomen.
The gut consists of foregut, midgut, and hindgut and extends the length of the body from anterior mouth to posterior anus. Foregut and hindgut are epidermal invaginations, being derived from the embryonic stomodeum and proctodeum respectively, and are lined by cuticle, as are all epidermal surfaces of arthropods. The midgut is endodermal and is responsible for most enzyme secretion, hydrolysis, and absorption.
The coelom is reduced to small spaces associated with the gonads and kidney. The functional body cavity is a spacious hemocoel divided by a horizontal diaphragm into a dorsal pericardial sinus and a much larger perivisceral sinus. Sometimes there is a small ventral perineural sinus surrounding the ventral nerve cord.
The hemal system includes a dorsal, contractile, tubular, ostiate heart that pumps blood to the hemocoel. Excretory organs vary with taxon and include Malpighian tubules, saccate nephridia, and nephrocytes. Respiratory organs also vary with taxon and include many types of gills, book lungs, and tracheae.
The nervous system consists of a dorsal, anterior brain of two or three pairs of ganglia, circumenteric connectives, and a paired ventral nerve cord with segmental ganglia and segmental peripheral nerves. Various degrees of condensation and cephalization are found in different taxa.
Development is derived with centrolecithal eggs and superficial cleavage. There is frequently a larva although development is direct in many. Juveniles pass through a series of instars separated by molts until reaching the adult size and reproductive condition. At this time molting and growth may cease or continue, depending on taxon.
Chelicerata sP
Chelicerata is a large taxon that includes spiders, scorpions, pseudoscorpions, ticks, mites, horseshoe crabs, sea spiders, and many others. The group originated in marine habitats but almost all modern chelicerates are terrestrial.
The body is divided into an anterior cephalothorax with six pairs of appendages and a posterior abdomen which, in most groups, does not bear appendages or has highly modified appendages. The first appendages of the cephalothorax are the chelicerae. Antennae are not present and the brain has two regions rather than the three found in mandibulates. Appendages are primitively biramous but are uniramous in almost all Recent taxa.
Euchelicerata
The segments of the cephalothorax are fused and covered by a dorsal shield, or carapace. Two median eyes are present.
Arachnida C
Arachnids are the terrestrial descendents of the early aquatic chelicerates. The taxon includes the mites, scorpions, pseudoscorpions, spiders, harvestmen, and several other taxa. All are adapted for a terrestrial existence with internal gas exchange and a waterproof integument. Some are capable of silk production and many use toxins to subdue or kill the prey. Most are carnivores, digestion is usually outside the body, and food is liquefied before ingestion.
Lipoctena
Lipoctena includes all arachnids except for scorpions. Book lungs, if present, are limited to two pairs. Lyriform organs are present in the ground plan.
Apulmonata
Apulmonata is the arachnids in which the ancestral book lungs have been lost.
Holotracheata
This taxon consists of arachnids with a single pair of spiracles opening into tube tracheae.
Cryptoperculata
Cryptoperculata includes the mites, harvestmen, and tick spiders. These arachnids have sensory second legs and the abdomen is broadly joined with the cephalothorax.
Opiliones O
Opiliones is characterized by having the abdomen broadly joined with the cephalothorax, a gonopore on the ventral cephalothorax, a penis or ovipositor, a segmented abdomen primitively with 10 segments, two median eyes on a prominent dorsal tubercle, and no lateral eyes. The egg-shaped body consists of cephalothorax and abdomen. The typical arachnid six-segmented cephalothorax is fused with the abdomen with no waist-like constriction marking the separation. Dorsally the cephalothorax is covered by an anterior carapace. The body is usually 5-10 mm in length but some are as small as 1 mm and others as large as 20 mm.
Opilione chelicerae are triarticulate and chelate, forming a pincer used to manipulate, kill, and dismember food. The pedipalps are leglike and usually have a sensory function although in some taxa (e.g. Laniatores) they are raptorial and adapted for grasping prey. The preoral cavity is enclosed anteriorly and posteriorly by the labrum and labium respectively and laterally by endites of the coxae of the pedipalps and first legs. The labrum and labium are midventral folds of the body wall just posterior to the base of the chelicerae and just posterior to the mouth respectively.
The most familiar taxa (i.e. Eupnoi) have long legs, typically exceedingly so and sometimes as much as 40 times the body length. The legs consist of coxa, trochanter, femur, patella, tibia, pretarsus (= metatarsus), and tibia. One or two (occasionally three) tarsal claws are present distally. The tarsi of derived Opiliones (e.g. Eupnoi) are typically secondarily divided into a maximum of 100 tarsomeres and the resulting multiarticulate tarsus is used as a very flexible whip or lasso to grasp slender substrata. True antennae being absent in chelicerates, most arachnids have replaced them functionally by modifying other appendages. In Opiliones the second legs are longer than the others and serve the functional role of antennae. Taxa with shorter legs may resemble spiders or mites.
Some abdominal segments are clearly visible and, in primitive taxa, are covered by 10 tergites. The anterior five abdominal tergites are frequently fused together to form a dorsal shield (= scutum) which is typically fused with the carapace. In some taxa the dorsal shield is present only in males. The posterior two tergites may be reduced or divided into two side-by-side, plates. The anterior abdominal sternites are displaced anteriorly to positions between some of the posterior legs. In the most familiar taxa (i.e. Eupnoi) they lie on the midline between the last three pairs of legs where they often form a genital operculum covering the gonopore. In most higher taxa the gonopore is on the midline posterior to the coxae of the third legs but in Eupnoi it is farther anterior, immediately posterior to the labium and thus just behind the mouth. Abdominal sternite 1 is lost and sternite 2 fuses with the cephalothoracic sternites.
The gut includes a pumping pharynx and extensive digestive cecae. The ancestral Malpighian tubules have been lost and excretion is the responsibility of a single pair of saccate nephridia (= coxal glands) and nephrocytes. Each nephridium opens via a nephridiopore on the posterior border of the third coxae. A pair of repugnatorial glands is present in the cephalothorax. These produce and release benzoquinones and phenols which are used to discourage predators. Poison glands and silk glands are absent. The heart is short, with two pairs of ostia and is in the anterior abdomen. Gas exchange is by a single pair of tubular tracheae and no book lungs are present. A pair of spiracles is located between the fourth coxa and abdomen. The nervous system is cephalized with all segmental ganglia coalesced to form a large subesophageal ganglion. Opiliones are gonochoric with direct sperm transfer using a penis. Females are equipped with an ovipositor. Both penis and ovipositor are unusual in arachnids. The gonopore is on the ventral cephalothorax, although in most arachnids it is abdominal. Unlike other arachnids, sperm transfer is direct, using the penis. Males are smaller than females, are more heavily sclerotized, and have larger chelicerae but sexual dimorphism is not pronounced. Males have longer legs and use them to find females.
Harvestmen are often called “daddy” (or granddaddy) longlegs” inviting confusion with similarly long-legged spiders in the family Pholcidae. This confusion has resulted in the widespread urban legend that harvestmen are the world’s most poisonous arachnids, when in fact they have no poison glands, do not bite, and are harmless. Most opiliones are predominantly predators feeding on small invertebrates but many, unexpectedly for an arachnid, consume vegetation, fungi, feces, and organic detritus. Also atypically for arachnids, harvestmen ingest particulate food.
Recent estimates of the number of described extant species vary between 5000-7000. New species are being described continuously.
Classification and Phylogeny
Giribet et al. (1999), using a combination of ribosomal DNA sequences and morphological characters, divided Opiliones O into two sister taxa, Cyphophthalmi sO and Phalangida sO. The name Phalangida, which has traditionally been used as a synonym of Opiliones (with ordinal rank) is now used for the sister taxon of Cyphophthalmi and includes all Opiliones except Cyphophthalmi, ie. Sironoidea SF (Sironidae F and Stylocellidae F). Phalangida sO is divided into three taxa, Eupnoi iO (Phalangioidea SF [Phalangiidae F and Leiobunidae F] and CaddoideaSF [Caddidae F]), Dyspnoi iO (Ischyropsalidoidea SF [Ischyropsalidae F] and Troguloidea SF [Dicranolasmatidae F and Nemastomatidae F]), and Laniatores iO (Travunioidea SF[Triaenonychidae F], Oncopodoidea SF [Oncopodidae F], and Gonyleptoidea SF [Phalangodidae F, Cosmetidae F, and Gonyleptidae F]) for a total of seven superfamilies and 11 families. The old taxon Palpatores which formerly included Eupnoi and Dyspnoi is now seen as paraphyletic and its two subtaxa, Eupnoi and Dyspnoi, are separated as being unrelated.
Cyphophthalmi sO includes the tiny “mite harvestmen” with short legs and bodies less than 3 mm in length. Their eyes are absent or indistinct and the repugnatorial glands open on a pair of cones protruding from the side of the cephalothorax. The legs are short and in general appearance they resemble mites. Tarsi are not divided into tarsomeres, thus consist of a single article. The carapace and first eight abdominal tergites are fused to form a sclerotized, dorsal shield (=scutum). The gonopore lacks an operculum. The segmented abdomen distinguishes them from mites. Mite harvestmen are found in leaf litter or caves and five species are known from North America north of Mexico.
Phalangida sO includes all the remaining Opiliones. Laniatores iO is a primarily tropical taxon but has representatives in the southern United States and Europe. About 100 species are known from the US. Members of this taxon have the coxae of the first three pairs of legs touching on the ventral midline, rather then being separated by a sternite. The integument is heavily sclerotized and some taxa have extraordinarily long legs. The pedipalps are raptorial with a claw. The leg tarsi are usually secondarily divided into tarsomeres. The gonopore is covered by an operculum which lies on the midline between the coxae of the fourth legs. Three families, Phalangodidae, Triaenonychidae, and Cosmetidae, have representatives in the US but most North American species, about 80, belong to Phalangodidae. Phalangodidae have two tarsal claws on the fourth leg. Triaenonychidae have only one tarsal claw on the fourth leg but it has teeth that make it appear to be trident. Sclerobunus robustus is a red species that lives in rotting logs in the Colorado mountains. Cosmetidae have three tarsal claws on the fourth legs, of which the median claw is smaller than the two laterals. The pedipalps are small with flattened, keeled articles. Vonones ornata is a small (4 mm), common, southeastern US cosmetid. Another family,Gonyleptidae, is restricted to the American tropics and includes the largest harvestmen (in terms of body size).
Eupnoi iO (formerly Palpatores, in part) includes two North American families, Phalangiidae and Caddidae. In Eupnoi (and Dyspnoi as well) a sternite separates the right and left coxae so they do not touch on the ventral midline. The primary spiracles are between the fourth coxae and the abdomen. Minute accessory spiracles on the tibia of the legs supplement the oxygen supply to the extraordinarily long legs. Coxa 2 has an endite. The tarsus of the pedipalp is longer than the tibia. Phalangiidae includes the common American and European harvestmen, with about 90 species known in North America. These are the common, familiar temperate zone harvestmen with relatively soft bodies and very long legs. Phalangiids move rapidly over the vegetation on which they live. Leiobunum, Phalangium, Mitopus, Odiellus, and Hadrobunus are important North American genera. Mitopus morio is a common European and North American species found in gardens and fields. The second article of the male chelicera has a dorsal projection. Opilio parietinus is another Holarctic species commonly found near human dwellings. Caddidae is characterized by enormous eyes. The tiny (2 mm) Caddo agilis occurs in eastern North America.
Dyspnoi iO (formerly Palpatores, in part) is represented in North America north of Mexico by four families, Trogulidae, Nemastomatidae, Ceratolasmatidae, and Sabaconidae. Dyspnoi are typically slow-moving soil dwellers with short legs whereas Eupnoi (viz Phalangiidae) are usually agile, fast, long-legged and live in vegetation. The tarsus of the pedipalp is shorter than the tibia. The coxae of the second legs lack endites. The gonopore is covered by an operculum. Trogulidae is characterized by a distinctive hood that projects anteriorly from the base of the eye tubercle to overhang the chelicerae. The dorsal shield (= scutum) includes the first five abdominal tergites. Soil particles adhere to the dorsum. The tarsi of the relatively short legs have 1-3 tarsomeres. Only one trogulid, Trogulus tricarinatus, occurs in North America and it, a predator of terrestrial snails, is an introduction from Europe. Nemastomatidae is represented by seven species north of Mexico. The legs are short, about the length of the body. Their pedipalps lack claws and the chelicerae are very short. The coxae of legs 1 and 4 have a row of marginal spines. The carapace and first five abdominal tergites are fused to form a dorsal shield. Nemastoma dasycnemum is a common eastern North American species. The tarsi have multiple tarsomeres. Many Nemastoma are cave dwellers. Collembola are a common prey. Ceratolasmatidae have sculpturing on the legs. Sabaconidae, with nine North American species (Sabacon andTaracus), have enormous chelicerae and pedipalps without claws. The legs are short.
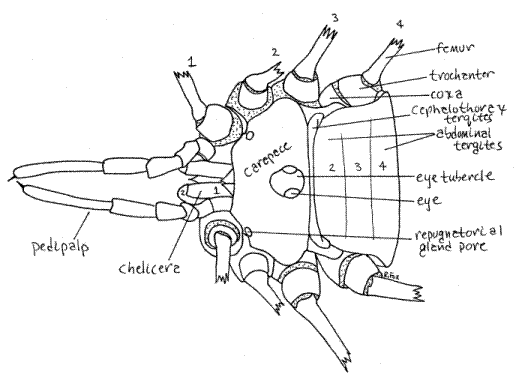
Figure 1. Dorsal view of an unidentified Phalangiidae from Greenwood, South Carolina. The legs have been removed. The chelicera articles and legs are numbered. Articular membranes are stippled. Opiliones30L.gif
Laboratory Specimens
Large harvestmen of several taxa are often common in most moist temperate zone localities, especially in summer and fall. Anatomical characteristics vary with taxon but this account should be usable with any Phalangida sO. It is written specifically for typical, long-legged phalangiids, such as Leiobunum vittatum, the striped harvestman, a common species in the eastern United States. The body of this species is a dull orangish color with a broad longitudinal black stripe along dorsal midline. The legs are black also. Leiobunum is a large genus with, for example, about 20 species in North Carolina and Tennessee.
Adult phalangiids, such as Leiobunum, appear, often in large numbers, in vegetation for a few weeks, often in late summer and fall, mate, oviposit, and die in winter. Most have a 1-year life cycle. Immature instars live near the soil surface, below the vegetation, where they are less likely to be noticed, thus giving the impression that the adults appear from nowhere in late summer. Adults may also undergo daily vertical migrations to different levels in the vegetation, including trees (where they are also less likely to be noticed). Extracting the long legs from the exuvium is a difficult maneuver for phalangiids and each leg must be pulled out individually using the chelicerae and pedipalps.
Specimens can be collected when available and preserved in 40% isopropanol until needed in the laboratory. Care must be taken to avoid separating the legs fro the body. Alternatively, if the season is appropriate, living specimens can be studied, either anesthetized or not. Anesthetization can be accomplished with carbon dioxide, ether, or chloroform. Smaller species can be found in leaf litter and extracted using a Berlese apparatus. This exercise is limited to external anatomy.
External Anatomy
Tagmata
As arachnids, harvestmen have a body divided into two tagmata, the anterior cephalothorax and the posterior abdomen, but the distinctions between them are obscure (Fig 1, 2, 3, 18-38). The anterior abdominal segments are fused with the cephalothorax so that their tergites are marked, at best, by faint transverse lines (Fig 1). There is no constriction between cephalothorax and abdomen.
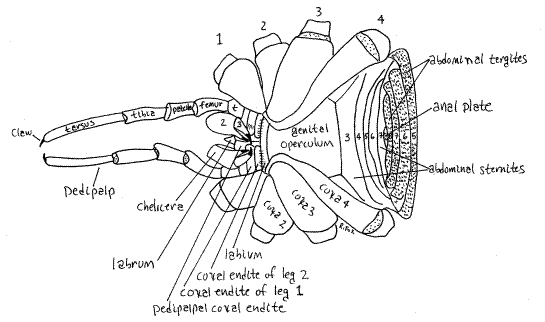
Figure 2. Ventral view of an unidentified Phalangiidae from Greenwood, South Carolina. The chelicera articles, legs, abdominal sternites, and abdominal tergites are numbered. Articular membranes are stippled. Abdominal tergites are shaded. Opiliones31L.gif
Cephalothorax
The cephalothorax is covered dorsally by a sclerotized, one-piece carapace consisting of the fused, indistinguishable tergites of the cephalothoracic segments. Somewhere on the dorsal midline, usually near the center of the carapace, is the eye tubercle bearing two median eyes, one on each side (Fig 1, 4, 18-38).
The ventral cephalothorax is covered by the large coxae of the four pairs of walking legs and the small labium (Fig 2, 18-38B). The anterior abdominal sternites are fused together to form the genital operculum which has been displaced far anteriorly between the leg coxae. The operculum covers the gonopore.
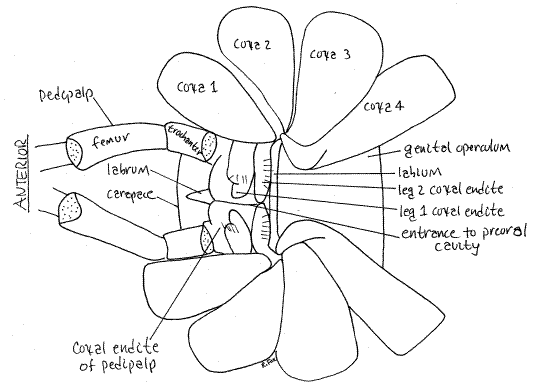
Figure 3. Ventral view of an unidentified Phalangiidae from Greenwood, South Carolina showing the appendages surrounding the preoral cavity. Opiliones33L.gif
Chelicerae
The anteriormost appendages are the two chelicerae (Fig 1,2,4, 18-38B). Opilione chelicerae are tri-articulate and the two distal articles form a prehensile chela.
Pedipalps
The second pair of appendages are the pedipalps (Fig 2,4, 18-38B). Although leglike, the pedipalps are not nearly so large as the walking legs. They consist of six articles which are, from proximal to distal, coxa, trochanter, femur, patella, tibia, and tarsus (Fig 4). In Eupnoi a tarsal claw is found at the tip of the undivided tarsus. The pedipalpal coxae have endites used to grind food. In some taxa the pedipalps have a large claw and are raptorial, used for capturing prey. The preoral cavity is bounded anteriorly by the conical, median labrum, laterally by thepedipalpal coxal endites, and posteriorly by the narrow, transverse labium (Fig 2, 3).
Figure 4. Lateral view of an unidentified Phalangiidae from Greenwood, South Carolina. The chelicera articles and legs are numbered. Articular membranes are stippled. Opiliones32L.gif
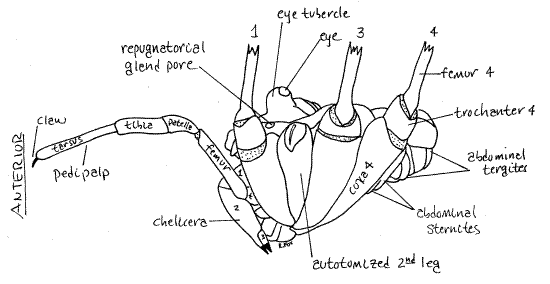
Figure 5. The right fourth leg of a harvestman in the family Phalangiidae. This leg is detached from the body and the coxa is not included. Opiliones29L.gif
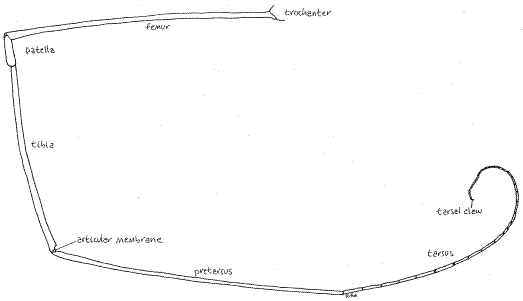
Legs
Harvestmen are famous for their exceedingly long legs (although some have short legs). Four pairs of walking legs are present on the thorax, of which the second are sensory and used as antennae. The second legs are the longest. The usual arachnid articles (coxa, trochanter, femur, patella, tibia, pretarsus, and tarsus) are present (Fig 5). One or two tarsal claws (rarely three) are present at the tip of the leg. The tarsi are divided into innumerable small secondary articles, or tarsomeres, to give the leg great flexibility. Phalangiids are capable of autotomy but jettisoned legs are not replaced (Fig 4). The fracture plane is between the coxa and trochanter.
Abdomen
The abdomen is segmented. Some anterior abdominal tergites may be fused to form a single sclerite, the dorsal shield (= scutum) , marked by faint transverse striae indicating the borders of the tergites (Fig 1). The dorsal shield is usually fused with the carapace. In many species the dorsal shield is present in males but not females. Some posterior abdominal segments remain separate and have individually visible abdominal tergites. A variable number of abdominal segments are represented ventrally by individual abdominal sternites (Fig 2). The number of free tergites and sternites varies with taxon.
A pair of spiracles is present laterally on the first abdominal segment between the fourth coxae and the abdomen. The anus is terminal in the center of the anal plate, which is the posteriormost tergite.
References
Bishop SC. 1949. The Phalangida (Opiliones) of New York. Proc. Rochester Acad. Sci. 9:159-235.
Edgar AL. 1966 . Phalangida of the Great Lakes region. Amer. Midland Naturalist 75:347-366.
Edgar AL. 1971. Studies on the biology and ecology of Michigan Phalangida (Opiliones). Misc. Publ. Mus. Zool. Univ. Michigan 144:1-64.
Eisner T. 2002. For love of insects. Belknap Press, Harvard University, Cambridge. 448 pp.
Giribet G, et al. 1999. Phylogeny of the arachnid order Opiliones (Arthropoda) inferred from a combined approach of complete 18S and partial 28S ribosomal DNA sequences and morphology. Molecular Phylogeny Evolution 11:296-307.
Giribet G, Boyer SL . 2002. A cladistic analysis of the cyphophthalmid genera (Opiliones, Cyphophthalmi). J. Arachnology 30:110-128.
Kaestner A. 1968. Invertebrate zoology, Vol II, Arthropod relatives, Chelicerata, Myriapoda. Wiley Interscience, New York. 472 pp.
Kury AB. Opiliones. http://acd.ufrj.br/mndi/Aracnologia/opiliones.html
Kury AB. Checklist of valid genera of Opiliones of the World. http://acd.ufrj.br/mndi/Aracnologia/checklaniator.htm
Levi HW, Levi LR. 2002. Spiders and their kin. St. Martin’s Press, New York. 160pp.
Macías-Ordóñez R. 2000. Touchy harvestmen. Natural History 109(8):58-61.
Piel W. Undated. Arachnology Home Page. Opiliones. www.arachnology.org/Arachnology/Pages/Opiliones.html
Ruppert EE, Fox RS, Barnes RB. 2004. Invertebrate Zoology, A functional evolutionary approach, 7 th ed. Brooks Cole Thomson, Belmont CA. 963 pp.
Savory T. 1977. Arachnida, 2 nd ed. Academic Press, New York. 340 pp.
Shear WA . 1980. A review of the Cyphophthalmi of the United States and Mexico, with a proposed reclassification of the suborder (Arachnida, Opiliones). Amer. Mus Novitates 2505:1-34.
Vanuytven H. Arachnology Home Page : www.arachnology.be/Arachnology.html
Weygoldt P. 1996. Chelicerata, pp. 449-497 in Westheide W, Reiger RM (Eds.) Spezielle Zoologie. Teil 1. G. Fischer Verlag, Stuttgart. 909 pp.
Supplies
Dissecting microscope
Living or preserved harvestman
Microdissecting tools