Invertebrate Anatomy OnLine
Diodora cayenensis ©
Keyhole Limpet
3jul2006
Copyright 2003 by
Richard Fox
Lander University
Preface
This is one of many exercises available from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises can be accessed by clicking on the links to the left. A glossary and chapters on supplies and laboratory techniques are also available. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Mollusca P, Eumollusca, Conchifera, Ganglionura, Rhacopoda, Gastropoda C, Prosobranchia sC, Vetigastropoda O, Fissurellidae F
(Fig 12-125)
Mollusca P
Mollusca, the second largest metazoan taxon, consists of Aplacophora, Polyplacophora, Monoplacophora, Gastropoda, Cephalopoda, Bivalvia, and Scaphopoda. The typical mollusc has a calcareous shell, muscular foot, head with mouth and sense organs, and a visceral mass containing most of the gut, the heart, gonads, and kidney. Dorsally the body wall is the mantle and a fold of this body wall forms and encloses that all important molluscan chamber, the mantle cavity. The mantle cavity is filled with water or air and in it are located the gill(s), anus, nephridiopore(s) and gonopore(s). The coelom is reduced to small spaces including the pericardial cavity containing the heart and the gonocoel containing the gonad.
The well-developed hemal system consists of the heart and vessels leading to a spacious hemocoel in which most of the viscera are located. The kidneys are large metanephridia. The central nervous system is cephalized and tetraneurous. There is a tendency to concentrate ganglia in the circumenteric nerve ring from which arise four major longitudinal nerve cords.
Molluscs may be either gonochoric or hermaphroditic. Spiral cleavage produces a veliger larva in many taxa unless it is suppressed in favor of direct development or another larva. Molluscs arose in the sea and most remain there but molluscs have also colonized freshwater and terrestrial habitats.
Eumollusca
Eumollusca is the sister taxon of Aplacophora and includes all molluscs other than aplacophorans. The eumolluscan gut has digestive ceca which are lacking in aplacophorans, the gut is coiled, and a complex radular musculature is present.
Conchifera
Conchifera, the sister taxon of Polyplacophora includes all Recent molluscs other than aplacophorans and chitons. The conchiferan shell consists of an outer proteinaceous periostracum underlain by calcareous layers and is a single piece (although in some it may appear to be divided into two valves). The mantle margins are divided into three folds.
Ganglioneura
Most Recent molluscs are ganglioneurans, only the small taxa Aplacophora, Polyplacophora, and Monoplacophora are excluded. Neuron cell bodies are localized in ganglia.
Rhacopoda
The mantle cavity is posterior in the ancestor although it may be secondarily moved anterior by torsion. This taxon includes gastropods and cephalopods.
Gastropoda C
Gastropoda is the largest molluscan taxon and is the sister group of Cephalopoda. Gastropods are united by descent from a torted ancestor although many exhibit various degrees of detorsion. Many are coiled and asymmetrical but the ancestor was probably symmetrical. Gastropods are relatively unspecialized molluscs known collectively as snails. The univalve shell, present in the ancestral gastropod and in most Recent species, is reduced or lost in many representatives. The flat creeping foot was inherited from their eumolluscan ancestors but gastropods have developed a distinct head with an abundance of sophisticated sense organs. The originally posterior mantle cavity has become anterior as a consequence of torsion, although detorsion has reversed this condition in many. Gastropods were originally gonochoric and most remain so but many derived taxa are hermaphroditic. Most are marine but many taxa have invaded freshwater and the only terrestrial molluscs are gastropods. Most have a single gill, atrium, and nephridium but the most primitive representatives have two of each. Only one gonad, the right, is present. The ancestor probably had an operculum. The nervous system is streptoneurous (twisted by torsion).
Prosobranchia sC
Prosobranchia was once one of three great gastropod subclasses but it is no longer considered to be a monophyletic taxon although the concept continues to be used as a pedagogical convenience. Prosobranchs are the gastropods most like the ancestral snails. They are torted and most have a shell and are coiled and asymmetrical. The mantle cavity is anterior. Most are gonochoric and most have an operculum. Most have only one gill in the mantle cavity but some primitive taxa have two. The right atrium is lost in most. Prosobranchs are specialized for life in marine benthic habitats although representatives are also found in freshwater and on land.
Vetigastropoda O
Vetigastropods, often known as archaeogastropods, have characteristics that in traditional phylogenies place them close to the ancestry of gastropods. The taxon includes keyhole limpets, slit snails, abalones, top and turban snails.
Zeugobranchia
Zeugobranchs have traditionally been considered to be the most primitive living gastropods (although there is some evidence that honor may belong to patellogastropods). In zeugobranchs we see many features expected of the ancestral gastropod. The mantle cavity is anterior as a consequence of torsion. Two gills are present and are bipectinate. Zeugobranchs are diotocardian, with two atria, and blood from each gill drains into its own atrium, right or left. The atria drain into a single median ventricle. The nephridia are paired (but the left is much smaller than the right and apparently has little or no function). A single large gonad releases its products through the duct of the right nephridium. The gonad is separated from the pericardial cavity.
The shell has a unique second aperture by which waste products are vented, presumably to avoid fouling the two gills. Surviving Zeugobranchia are the slit snails, abalones, and keyhole limpets. All are torted but may or may not be coiled as adults.
Fissurellidae F
Keyhole limpets are symmetrical and uncoiled, although this condition is secondary, not primary. The shell is equipped with an anal pore or anal slit.
Laboratory Specimens
Any species of Fissurellidae can be used with this account of the anatomy of Diodora cayenensis but it will apply best to species of Diodora. Diodora cayenensis is a common species occurring from Maryland to Brazil on the American Atlantic coast. Diodora aspera occurs in shallow water from Alaska to Baja California on the Pacific Coast. Megathura crenulata is a large keyhole limpet found in southern and Baja California. Diodora apertura occurs on the west coast of Britain. Diodora dysoni occurs in south Florida and the Caribbean south to Brazil.
As zeugobranchs, keyhole limpets are good examples of the predicted ancestral gastropod condition with respect to the atria, nephridia, gills, and the pattern of water circulation through the mantle cavity. Furthermore, keyhole limpets are symmetrical and uncoiled. Most of the important features illustrating the zeugobranch condition can be studied without opening the body of the animal. It is necessary to remove the shell and open the mantle cavity but further dissection is not required. Interested students are encouraged, as always, to proceed beyond the written instructions and may investigate the internal anatomy if they wish and time permits.
Living specimens should be relaxed and dissected in isotonic magnesium chloride. Preserved material should be dissected submersed in tapwater. The dissection should be conducted in a small dissecting pan on the stage of a dissecting microscope.
External Anatomy
" Remove the animal from its shell using a scalpel to scrape (not cut) the soft tissue away from its attachment to the shell. Insert the blade of the scalpel between the body and the shell and work it completely around the circumference of the shell. Keep the blade in contact with the shell at all times and do not cut any tissues. The horseshoe-shaped attachment (insertion) of the pedal retractor muscle adheres tightly to the shell (in living specimens) and is hardest part to free.
Shell
The shell may be studied attached to the animal or separately. The shell of keyhole limpets is a single piece shaped like a broad oval cone, resembling a volcano, complete with a crater at the summit (Fig 1). The shell is not coiled and is bilaterally symmetrical (Fig 12-22A). While this symmetrical condition resembles the hypothesized condition of the shell of the ancient prosobranch ancestor, in fissurellids it is not primitive. Keyhole limpets arise from ancestors with coiled shells and, indeed, each individual begins life with an asymmetrical coiled shell but loses all but the large body whorl during development.
Figure 1. Dorsal and side views of the shell of a keyhole limpet, Diodora. Gastrop150La.gif
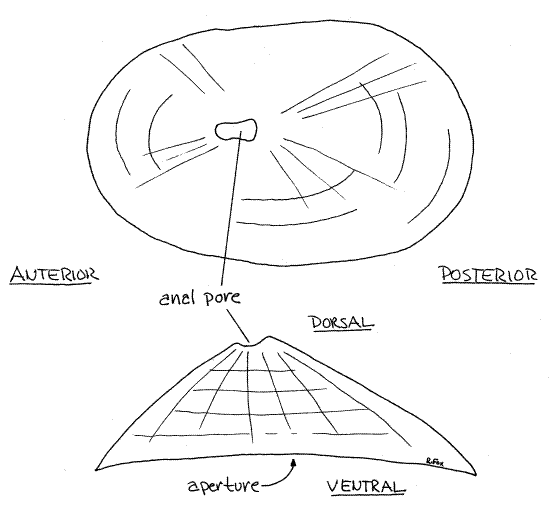
Examine the interior of the shell. Note the lustrous, smooth, nacreous innermost layer. It is secreted by the entire dorsal surface of the mantle. Look for the horseshoe-shaped pedal retractor muscle scar. It is the insertion site of the pedal retractor muscle but is indistinct and you may not find it.
The shell forms a shield over the dorsal surface of the animal but does not provide a commodious retreat as do the shells of higher gastropods. Unlike the shells of all other gastropods, those of keyhole limpets have two openings. The broad open base of the cone is the large aperture from which the foot and head protrude. It is homologous to the aperture of other gastropod shells.
The second opening is the much smaller anal pore situated at the apex, or summit, of the cone (Fig 1). It is the "crater" of the “volcano” mentioned above. In Diodora the anal pore is displaced toward the anterior end and is not in the center of the shell. The presence of the anal pore distinguishes the keyhole limpets from the several other (unrelated) kinds of limpets and is responsible for the common name "keyhole". The anal pore is the adult remnant of a large marginal anal slit in the shell of the developing juvenile (Fig 12-26).
The outside of the shell is variously ornamented with radial and concentric ridges, scales, beads, or spines depending on species (Fig 1). The interior is smooth and glossy.
Diodora is bilaterally symmetrical and the adult shows no signs of coiling, although it is torted. The visceral mass and mantle cavity are rotated 180 ° counterclockwise with respect to the foot, thus placing the mantle cavity and anus at the anterior end and identifying the limpet as a gastropod.
Foot
Examine the body of the animal. The dorsal body wall (mantle) is very thin and transparent so that most of the organs in the visceral mass can be seen without further dissection or, at the most, by opening the mantle cavity.
The body consists of the foot, visceral mass, and head. The large, muscular, oval foot occupies the entire ventral surface (Fig 2). It is broad and flat and is used for adhering to and creeping over hard substrata.
The foot normally extends below and outside the margins of the shell but when threatened the shell can be pulled ventrally to contact the substratum and protect the soft parts, including the foot.
The foot is attached to the ventral surface of the shell by a large U-shaped (in frontal section) pedal retractor muscle (Fig 2) derived from the separate right and left pedal retractor muscles of ancestral gastropods (Fig 12-26). The open end of the "U" faces anteriorly. When this muscle contracts, it pulls the shell toward the foot, which is normally attached to the substratum, so the shell is thus pulled against the substratum.
The columellar muscle of coiled gastropods is derived from the pedal retractors. In coiled gastropods only the right pedal retractor muscle is present, as the columellar muscles, and it is responsible for withdrawing the foot into the aperture of the shell. It extends from the central axis of the coiled shell (the columella) to the foot. This is essentially the same action, origin, and insertion as the pedal retractor muscle of keyhole limpets.
A longitudinal row of small papillae, the epipodial tentacles. Extends along the dorsal edge of the foot on each side of the body. It has been claimed, but not substantiated, that these are repugnatorial but their innervation suggests a sensory function.
Head
Anchor the head and posterior end of the foot to the wax of the dissecting pan using a #l insect pin at each end.
The head is located dorsal to the foot at the anterior end of the visceral mass (Fig 2, 12-22B). The visceral mass contains most of the visceral organs and occupies most of the interior of the shell. It is situated above the foot and is always covered by the shell. It will be studied in more detail later.
A cylindrical, tubular snout is directed ventrally and slightly anteriorly from the head. The mouth is located at the center of the distal end of the snout.
A pair of cephalic tentacles extends from the dorsolateral corners of the head (Fig 2). Each bears a lateral eye at its base.
The anterior-most right epipodial tentacle is immediately posterior to the right cephalic tentacle. It closely resembles, in shape and position, the penis of higher prosobranch gastropods and has been mistaken for than organ. It is present in both sexes, however, and appears to have a sensory, rather than a reproductive, role.
Mantle and Mantle Cavity
The mantle is the dorsal body wall of the visceral mass. It is attached to the inside of the shell above the visceral mass. Peripherally the mantle protrudes from beneath the shell as a prominent fold of the body wall, called the mantle skirt. The skirt forms a collar around the aperture. It is formed into three folds and is homologous to the enormous mantle skirt of bivalves, which also bears three folds (usually).
The outer fold is the smallest. It lies against the edge of the shell and its lateral, or outer, surface secretes the middle, prismatic, layer of the shell. The medial surface of the outer fold is thought to secrete the outer shell layer, or periostracum. The innermost, lamellar, nacreous shell layer is secreted by the entire outer surface of the mantle and not by a fold. Shell secretion in gastropods has not been carefully studied and the much of our knowledge of the subject is based on extrapolations what is known of bivalves.
The middle mantle fold is sensory and bears numerous mantle tentacles (Fig 2). The medial, or inner, fold of the mantle margin is the largest and is muscular. Muscles arising in this fold insert on the inner surface of the shell near the margin of the aperture. These muscles attach the mantle to the shell.
Anteriorly the mantle overhangs a deep, median recess, the mantle cavity proper. This cavity is open to the sea via a large anterior opening above the head. The cavity itself lies above the anterior visceral mass and under the anterior shell. Use a probe or needle to lift the mantle skirt and demonstrate the presence of the mantle cavity.
Laterally the mantle skirt overhangs a longitudinal channel called the pallial groove (pallial = mantle) on the side of the animal between the foot and the visceral mass. It is a lateral extension of the anterior mantle cavity but by no means as deep. The epipodial tentacles, arrayed in a longitudinal row, protrude into the pallial groove. They arise from the lateral body wall immediately dorsal to the foot. Do not confuse the epipodial tentacles with the mantle tentacles of the middle mantle fold.
Push the head ventrally and look into the anterior mantle cavity again. The most conspicuous structures in it are the two bipectinate gills (Fig 2, 12-22B). The presence of two gills is one of the reasons for studying this animal. Among all prosobranch gastropods it is only zeugobranch vetigastropods that have two gills, the presumed ancestral condition. In all others the right gill is lost.
The anterior ends of the horseshoe-shaped pedal retractor muscle form the lateral walls of the mantle cavity (Fig 2).
Find the dorsal opening in the posterior roof of the mantle cavity and note the short sensory tentacles around it. This is the exhalant aperture from the mantle cavity (Fig 2). Its position coincides with that of the anal pore at the summit of the shell.
" Open the mantle cavity with a median, dorsal, longitudinal incision through its roof. Use fine scissors and begin the incision at the anterior edge of the mantle, above the head, and cut posteriorly to the exhalant aperture. Deflect and pin the roof of the mantle cavity and examine its interior.
Figure 2. Dorsal view of Diodora dysoni from Big Pine Key, Florida. The shell has been removed and mantle cavity has been opened with a median incision and its two sides deflected. Gastrop151La.gif
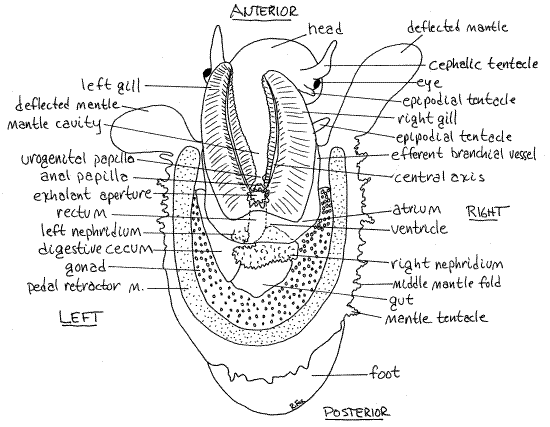
The two large, triangular, bipectinate gills fill most of the mantle cavity (Fig. 2, 12-22B). The efferent membrane attaches them by their narrow bases and part of their lateral (efferent) borders to the floor of the mantle cavity. Their tapered apices protrude anteriorly out of the mantle cavity.
Each gill has a central axis from which arise two rows of sheetlike filaments, one on each side of the axis. It is the presence of two rows of filaments on each gill that makes these gillsbipectinate. Bipectinate gills are predicted for the ancestral gastropod. The gills of higher prosobranchs (mesogastropods and neogastropods) are monopectinate with only one row of filaments. An inconspicuous osphradium is located along the medial side of the base of the efferent membrane, parallel to and beside the gill axis.
There is a pair of hypobranchial glands in the posterior roof of the mantle cavity, above the bases of the gills.
The anus opens into the posterior end of the mantle cavity on top of a short anal papilla between the bases of the gills (Fig 2, 12-22B). The similar urogenital papilla is located to the right and slightly ventral to the anal papilla (Fig 2, 12-22B). The gonad and right kidney open via the urogenital pore at the end of this papilla. The vestigial left kidney opens via a tiny inconspicuous pore located to the left of the anal papilla (Fig 12-22B). It is difficult to find. The anus and urogenital papillae are is immediately ventral to the exhalant aperture and anal pore.
While looking into the open mantle cavity, mentally trace the flow of water through it (Fig 12-22B). Water enters anterolaterally, on both sides of the head, flows posteriorly and medially over the osphradia and gill filaments, over the nephridiopore, and over the anus. It then turns dorsally to exit via the exhalant aperture and anal pore. This arrangement allows water to enter on both sides of the head and flow over both gills and the anus without running the risk of fouling the gills or the mouth with feces, urine, or gametes. It requires, of course, the presence of an anal pore for the exhalant current.
In higher vetigastropods, mesogastropods, and neogastropods there is no anal pore or its equivalent and the respiratory water current must enter and leave anteriorly. Only the left gill is present in the higher snails and water enters the left anterior side of the mantle cavity, flows over this gill, then over the anus on the right side of the mantle cavity and then out the right anterior side of the cavity.
Hemal System
Look at the roof of the visceral mass posterior to the mantle cavity. The transparent, colorless pericardial cavity is located immediately posterior to the junction of the bases of the two gills (Fig 2). In it is located the diotocardian heart. This organ consists of a single central ventricle and two lateral atria.
A swollen, transparent atrium is located on the postero-lateral corner of each gill, beside the median ventricle. A narrow efferent branchial vessel lies along the lateral border of each gill (Fig 2). Oxygenated blood drains from the lateral edge of the gill into this vessel. The vessel in turn empties into its atrium which opens to the ventricle. Contractions of the ventricle force oxygenated blood into the aorta to the body.
Excretory System
Although right and left metanephridia, or kidneys, are present, the right is much larger than the left (Fig 2). The right nephridium is a pale yellow (in life), folded, saccate organ lying immediately posterior to, and partly covered by, the ventricle. Its lumen is connected to the pericardial cavity by a renopericardial canal, which you will probably not see. The right nephridium opens to the exterior by a nephridiopore on the tip of the urogenital papilla.
The left nephridium is present but it is small and vestigial. It is a simple, ciliated sac that opens into the posterior mantle cavity via a tiny nephridiopore on the posterior wall of the mantle cavity to the left of the anus. It has no connection with the pericardial cavity and its function is not known.
The right nephridium is lost in higher prosobranchs and in them the left nephridium persists and is important as the only complete functional nephridium. All that remains of the right nephridium of higher gastropods is a small region that contributes to the genital duct.
Digestive System
Most of the digestive system is hidden from view by other organs lying over it and cannot be seen without opening the body. There follows a brief account of the anatomy of the gut but it is not intended as dissection instructions and you should not attempt to open the gut or visceral mass at this time. After you have completed the study of externally visible anatomy you may return to this section as use it as a guide to help you find the most conspicuous internal features if you wish. At present find only the structures in bold type. They are visible from the exterior without further dissection.
The ventral mouth opens into a small buccal cavity. The radula lies in a pouch, the radular sac, off the floor of the buccal cavity. The esophagus extends posteriorly to the stomach on the left side of the visceral mass. The walls of the esophagus are expanded to form a large glandular region. The stomach is pyriform with its swollen end posterior.
The style sac is the narrow anterior end of the stomach and may be visible at the surface of the visceral mass to the left of the nephridium and ventricle. The style sac contains a rotating mass of feces and mucus (the protostyle) but no crystalline style. The stomach conforms with the prediction for the ancestral mollusc except for the effects of torsion. Because of torsion, the esophagus enters the bulbous posterior end of the stomach and the intestine exits the narrow anterior end. Torsion bends the gut into a "U" at the level of the stomach and reverses the customary relative positions of the esophagus and intestine.
No gastric cecum is associated with the stomach. There is a chitinous gastric shield in the posterior stomach and two ducts from the digestive ceca open here. The stomach walls have a sorting field of ciliated ridges and grooves. Two typhlosoles border a ciliated intestinal groove that extends along the style sac into the intestine.
The intestine exits the style sac at the narrow anterior end of the stomach and makes a single loop to end up on the midline as the rectum (Fig 2). This terminal region of the intestine is visible on the dorsal midline where it passes through the pericardial cavity and ventricle to end at the anal papilla and anus.
The mollusc intestine is minimally involved in digestion and absorption and its major role is compaction of the feces into pellets. A long intestine is usually associated with the need to package the feces into pellets that will not disintegrate in the mantle cavity and foul the gills or mouth. The intestine of Diodora is short and does not make fecal pellets. The presence of the anal pore in the fissurellids minimizes the danger of fouling the gills so feces do not need to be tightly packaged. The intestine of true limpets (e.g. Pectura), which lack the apical aperture, is much longer and manufactures fecal pellets.
A "siphon" similar to that of echiurans and echinoids parallels the intestine. It is a small-diameter ciliated tube that diverges from the anterior intestine just below the style sac and then rejoins it near the anus. It is formed by the closing of the intestinal groove but its function is unknown. In other invertebrates with such a structure its function is to shunt large amounts of inadvertently ingested water past the stomach and avoid thereby dilution of digestive enzymes.
Portions of the two large, dark digestive ceca can be seen on the surface of the visceral mass (Fig 2). Combined, the ceca are the largest organ in the visceral mass and the most conspicuous organ on the surface of the mass. Each cecum is connected to the stomach via its own duct. The ceca are hollow and composed of an abundance of tiny branching tubules that eventually connect with the stomach.
The digestive ceca secrete hydrolytic enzymes and release them into the stomach where extracellular digestion begins. The products of hydrolysis and particulates move into the digestive ceca where extracellular digestion continues. The products of digestion are absorbed by the epithelium of the ceca. In zeugobranchs there is also some phagocytosis and intracellular digestion in the digestive ceca. The digestive ceca is surrounded by the hemocoel and immersed in blood.
>1a. Fissurellids are browsers on hard substrata and the British species, Diodora apertura, feeds on sponges (Hymeniacidon, Halichondria). You may wish to prepare a wetmount of the contents of the rectum or stomach and examine it for sponge spicules. Poll the class to see if anyone has found evidence of feeding on sponges. Look for other recognizable food items in the gut contents. <
Reproductive System
Fissurellids are gonochoric and a single, large gonad, either ovary or testis occupies much of the space in the ventral visceral mass (Fig 2). Portions of it are visible on the dorsal surface of the mass especially during periods of reproductive activity. This single gonad is thought to be homologous to the right gonad of the ancestral mollusc.
The gonad is not intermingled with the digestive ceca. The gonad empties to the exterior via the lumen of the right nephridium. The duct from the gonad opens into the renopericardial canal between the pericardial cavity and the right nephridium (Fig 12-26). Gametes pass through the canal into the lumen of the nephridium and then exit into the mantle cavity via the urogenital pore on the urogenital papilla.
Fertilization is external and there is none of the complex elaboration of the genital duct associated with internal fertilization seen in mesogastropods and neogastropods.
>1b. Determine the sex of your specimen by examining the gonad. Eggs are clearly evident and recognizable in females. Prepare a wetmount of a small fragment of the gonad in seawater. Look for gametes. Eggs will be large nucleated spheres. Sperm will be tiny and flagellated. If activated by the seawater they may be swimming. <
The claim has been made, on the basis of correlations between sex and body length, that Diodora, although usually gonochoric, is sometimes a consecutive hermaphrodite. If this is true, any individual you examine will be either male or female, but its sex will change during its life. Diodora, if it is hermaphroditc, is protandric, meaning that each individual begins life as a male, then changes to female as it grows to its final size.
Protandry is the more common of the two forms of consecutive hermaphroditism because it allows the animal to make sperm, which are relatively inexpensive, while it is small and eggs, which require more resources, when it is large. The alternative, female first then male, is protogyny and is less common.
>1b. Measure the length of the shell of your specimen (in mm) and determine its sex if you have not already done so. Record these data on the chalkboard along with similar data from other students. Examine the compiled data and see if they reveal a tendency for Diodora to be male when small and female when large. Do you think your species is hermaphroditic? Do not place too much faith in conclusions based on small samples. <
Development is direct, there is no free-living veliger, and the young emerge from the large yolky eggs as miniature adults.
Internal Anatomy
" If you wish to see the digestive system in its entirety, you may dissect the visceral mass. First remove the very thin, transparent body wall from its dorsal surface.
Take this opportunity to lift gently the superficial organs so you can see those lying deep to them. Examine the extent of the digestive gland and the gonad and confirm the earlier statement that they are much larger than their dorsally visible portions.
Trace the digestive system by opening it and following its lumen through the surrounding tissues. This is the best way to trace the gut in almost any mollusc. Insert one point of your finest scissors into the mouth and make a median dorsal incision through the dorsal body wall to open the gut lumen. Follow the gut posteriorly by opening it in this manner. Find the regions described in the digestive system section.
Behavior
>1d. At least one species, Diodora aspersa, responds in an interesting and predictable manner to predatory asteroids (forcipitulates), or to water formerly inhabited by these sea stars. In response to a kairomone from the star, the snail extends the mantle to cover most of its shell. The result is that the star’s tube feet cannot obtain the necessary grip on the shell to pull the limpet from its rock. There is some indication that the mantle surface may be repugnant to the star (Margolin, l964).
If living limpets and sea stars or other echinoderms are present in your laboratory, make some simple tests to see if your limpet species will respond to any of the echinoderms available. Record the nature of the response and the combinations of limpet/echinoderm species that elicit it. <
>1e. Watch a living limpet crawl across a culture dish of seawater. Squirt a little carmine/seawater in front of the snail and see if you can visualize the respiratory current. <
>1f. Turn a living limpet on its back in a dish of seawater and see if it can turn itself back over. Limpets are unusual among gastropods in their inability to right themselves. <
>1g. Place a living limpet with its foot down on a clean microscope slide in a dish of seawater. The animal will quickly attach to the slide, which can then be turned over and supported above the bottom of the dish by short lengths of thick glass rod or by other means. Examine the foot and head of the limpet with the dissecting microscope. Find the mouth and watch the operation of the radula as it is repeatedly protruded and retracted (Fig 12-2). <
>1h. Use a small paintbrush to apply a coat of India ink to the surface of a small glass plate or microscope slide. Let the ink dry thoroughly. Place the plate in a dish of seawater (but not in your aquarium) and put a limpet on it. Let the limpet attach and crawl over the surface of the plate. With a little luck, the limpet will scrape the surface of the slide with its radula as it crawls across it. The radula will remove the ink in a pattern characteristic of the species. Remove the limpet from the glass, remove the glass from the dish, and gently rinse the seawater from it. Let it dry. The plate can now be painted or sprayed with shellac or varnish and retained as a permanent record of its species specific radular pattern. You may want to try this with other microphagously browsing snails or chitons and compare the patterns. <
References
Abbott DP. l987. Observing Marine Invertebrates. Stanford, Palo Alto, 380 p. (Megatebennus bimaculatus and Fissurella volcano)
Fretter V, Graham A. l994. British Prosobranch Molluscs, 2 nd ed. Vol 161. Ray Society, London. 820pp.
Hyman LH. 1967. The Invertebrates, IV. Mollusca. McGraw-Hill, New York. 792p.
Margolin AS . l964. The mantle response of Diodora aspersa. Animal Behavior l2:l87-l94.
Ruppert EE, Fox RS, Barnes RB. 2004. Invertebrate Zoology, A functional evolutionary approach, 7 th ed. Brooks Cole Thomson, Belmont CA. 963 pp.
Supplies
Dissecting microscope
Dissecting set with microdissecting tools
Diodora or other fissurellid, living or preserved
Small dissecting pan
#1 stainless steel insect pins:
Isotonic magnesium chloride for living specimens
microscope slides
seawater for living specimens
carmine/seawater suspension