Invertebrate Anatomy OnLine
Calpodes ethlius Larva ©
Greater Canna Leafroller
30jun2006
Copyright 2001 by
Richard Fox
Lander University
Preface
This is one of many exercises available from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises can be accessed by clicking on the links to the left. A glossary and chapters on supplies and laboratory techniques are also available through this link. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Arthropoda P, Mandibulata, Tracheata, Hexapoda SC, Insecta C, Dicondylia, Pterygota, Metapterygota, Neoptera, Eumetabola, Holometabola, LepidopteraO, Hesperioidea SF, Hesperiidae F, Hesperiinae sF (Fig 16-15, 20-14, 20-15, 21-23)
Arthropoda P
Arthropoda, by far the largest and most diverse animal taxon, includes chelicerates, insects, myriapods, and crustaceans as well as many extinct taxa. The body is segmented and primitively bears a pair of jointed appendages on each segment. The epidermis secretes a complex cuticular exoskeleton which must be molted to permit increase in size. Extant arthropods exhibit regional specialization in the structure and function of segments and appendages. The body is typically divided into a head and trunk, of which the trunk is often itself divided into thorax and abdomen.
The gut consists of foregut, midgut, and hindgut and extends the length of the body from anterior mouth to posterior anus. Foregut and hindgut are epidermal invaginations, being derived from the embryonic stomodeum and proctodeum respectively, and are lined by cuticle, as are all epidermal surfaces. The midgut is endodermal and is responsible for most enzyme secretion, hydrolysis, and absorption.
The coelom is reduced to small spaces associated with the gonads and kidney. The functional body cavity is a spacious hemocoel divided by a horizontal diaphragm into a dorsal pericardial sinus and a much larger perivisceral sinus. Sometimes there is a small ventral perineural sinus surrounding the ventral nerve cord.
The hemal system includes a dorsal, contractile, tubular, ostiate heart that pumps blood to and from the hemocoel. Excretory organs vary with taxon and include Malpighian tubules, saccate nephridia, and nephrocytes. Respiratory organs also vary with taxon and include many types of gills, book lungs, and tracheae.
The nervous system consists of a dorsal, anterior brain of two or three pairs of ganglia, circumenteric connectives, and a paired ventral nerve cord with segmental ganglia and segmental peripheral nerves. Various degrees of condensation and cephalization are found in different taxa.
Development is derived with centrolecithal eggs and superficial cleavage. There is frequently a larva although development is direct in many. Juveniles pass through a series of instars separated by molts until reaching the adult size and reproductive condition. At this time molting and growth may cease or continue, depending on taxon.
Mandibulata
Mandibulata includes arthropods in which the third head segment bears a pair of mandibles. As currently conceived this taxon includes myriapods, hexapods, and crustaceans. Appendages may be uni- or biramous and habitats include marine, freshwater, terrestrial, and aerial.
Tracheata
Myriapods and hexapods share tracheae and a single pair of antennae and are sister taxa in Tracheata. Crustaceans, which have gills and lack tracheae, are excluded and form the sister group.
Hexapoda
The body is divided into three tagmata; head, thorax, and abdomen. Appendages are uniramous and a single pair of antennae is present. Three pairs of legs and two pairs of wings are found on the thorax of most adults. Hexapod legs are uniramous although there is increasing evidence that they evolved from multiramous appendages of their ancestors. Gas exchange is accomplished by trachea. Excretory organs are Malpighian tubules and the end product of nitrogen metabolism is uric acid. There is relatively little cephalization of the nervous system. Insects are gonochoric with copulation and internal fertilization.
Insecta C
Most hexapods are insects. A few hexapod taxa (orders) lack wings and have primitive mouthparts recessed into the head and belong to Entognatha, the sister taxon of Insecta. Insects have ectognath mouthparts and the adults (imagoes) of most taxa have wings.
Pterygota
The winged insects. These insects are derived from a winged common ancestor. Adults of most taxa have wings although they have been lost in some.
Eumetabola
Juveniles have no ocelli and there are six or fewer Malpighian tubules.
Holometabola
The final larval instar pupates and undergoes a radical metamorphosis in which it is converted to an imago, or adult. The imago is sexually mature and in most taxa has wings whereas larvae are immature and wingless. During metamorphosis many or most larval tissues are dismantled and adult structures built anew. Wings, for example, are manufactured from clusters of undifferentiated cells known as imaginal discs but not from preformed wingpads as in pauro- and hemimetabolous insects.
Lepidoptera O
Butterflies and moths. Order consisting of the polyphyletic “moths”, skippers (Hesperoidea), and scudders (Papilionoidea). Skippers and scudders are together known as “ butterflies”. Wings, body, appendages covered with pigmented, dust-like epidermal scales or hair-like setae. Adults with highly derived sucking mouthparts and liquid diet. Adult mouthparts consist of large labial palps and a coiled tubular proboscis derived from the maxillary galeae. Mandibles absent. Larvae herbivorous with typical chewing mouthparts. Larval labial glands modified as silk glands. Holometabolous complete metamorphosis in a pupal stage. In many moths pupation occurs in a silk cocoon but most butterflies have no cocoon and the pupa is known as a chrysalis, sometimes associated with a few silk fibers. Lepidopteran pupae are obtect, with the appendages attached to the body over their entire length.
Introduction
Several species of Lepidoptera have larvae large enough to be conveniently dissected as an introduction to caterpillar anatomy. Chief among them are the Tomato and Tobacco Hornworms in the genus Manduca (Sphingidae) but Calpodes ethlius (= Hesperia ethlius), the Greater Canna Leafroller, should be considered for this purpose in places and seasons where and when it is available alive. It has the advantage of having a transparent body wall through which can be observed most of the internal organs in a living, active animal.
Two species of leaf-rolling neotropical caterpillars occur on ornamental cannas (Canna) in the southern United States. One, the Lesser Canna Leafroller, Geshna cannalis (Pyralidae), is currently known only from the southeastern US but is presumed to occur in tropical America since its host plants (cannas) have a tropical distribution. It is a bit small for use in the teaching laboratory.
The second, the much larger larva of which is the subject of this exercise, is the Greater Canna Leafroller, Calpodes ethlius (Hesperiidae) known from the Caribbean and much of South America including the Galapagos Islands and also Bermuda. It has been introduced into the southern United States where it feeds on ornamental cannas. The adult is a butterfly known as the Canna Skipper or Brazilian Skipper. It is largely restricted to members of the plant family Cannaceae but also occurs on the closely related arrowroot (Maranta arundinacea), which is grown commercially as a source of culinary arrowroot.
Leafroller larvae roll the leaf margins of the host plant to form a tube held together by silk and used as a domicile offering protection from predators and pesticides. Eventually the domicile and nearby leaf tissue is consumed and another constructed near a fresh food supply. Unlike the Lesser Canna Leafroller, Calpodes larvae flick their frass (fecal pellets and debris) away from the domicile and do not allow it to accumulate, presumably avoiding thereby attracting predators or parasitoids. The fecal pellets are flipped away by the anal comb in a process known as “fecal firing”. Adult Greater Canna Leafrollers are relatively nondescript, fast-flying, brown butterflies. Females, which are slightly larger than males reach 27 mm in length.
Greater Canna Leafrollers are reported to be most common on red-leaved cannas but also occur on those with green leaves. In the South Carolina piedmont, I have found the caterpillars on yellow-flowering cannas and Bengal Tiger Cannas (orange flowers), both of which have green leaves. Calpodes is host to several species of parasitoid chalicidid, trichogrammatid, and encyrtid wasps as well as some tachinid flies (McAuslane & King, 2000).
The family Hesperiidae consists of about 3500 species of strong fliers known as skippers for their irregular “skipping” flight pattern. The larvae usually inhabit domiciles constructed of leaves held together by silk secreted by the salivary glands. Larvae have a characteristic appearance with a large head with a narrowed neck and are easily recognized by this trait.
Like that of most butterflies, the life cycle consists of egg, five larval instars, pupa, and imago. Butterflies are holometabolous insects in which the body undergoes a radical metamorphosis during the pupal instar. During this metamorphosis wings develop de novo from imaginal discs (clusters of undifferentiated embryonic cells) in the larval thorax and muscles are reorganized for flight. The mouthparts are converted from the primitive chewing design suited for feeding on plant tissue to highly derived sucking mouthparts adapted for ingesting liquids. The gut is reorganized to accommodate differences in larval and adult diets. The antennae form anew from imaginal discs in the head and the reproductive system matures. The prolegs are completely lost and the adult compound eye develops near the old larval ommatidia. The nervous system is reorganized, in part through condensation and cephalization of ganglia.
The fifth instar reaches lengths of 65 mm before pupating, making it more than large enough for easy dissection. The larvae are smooth-bodied with little external ornamentation and are tapered at both ends, being widest in the middle. They are naked, with scattered, inconspicuous setae. Fifth instar larvae have an orange head capsule although that of earlier instars is black. The larva pupates in its leaf nest as a bright green pupa. The pupa has a black anterior spine, a very large proboscis extending posteriorly beyond the tip of the abdomen, and is about 35 mm in length. It’s rudimentary silken cocoon consists a platform of a few threads. Photographs of all life history stages, as well as a discussion of biology, can be found at McAuslane & King (2000).
This exercise is written for use with fifth instar living or freshly sacrificed caterpillars. It can also be used with other instars and with preserved specimens although color and texture descriptions are not applicable to preserved material. If possible the specimens should be freshly collected so the gut is filled with ingested leaf particles.
External Anatomy
Examine a living, active (i.e. unanesthetized) late instar larva of the Canna Skipper, Calpodes ethlius. Most of the external anatomy is best studied using an active animal and, because of the transparent body wall, much of the internal anatomy is visible and can be studied prior to (or without) dissection. The beating heart, pericardial sinus, perivisceral sinus, tracheae, Malpighian tubules, salivary (silk) glands, and the enormous gut and its contents can be seen through the body wall of living specimens. With a little patience, the external anatomy can be studied without anesthetizing the specimen and it is desirable to do so to take advantage of the opportunity to observe motion and behavior. Sometimes the caterpillar will sit quietly on the tip of your index finger, which can then be oriented under the lens of a dissecting microscope to provide the desired view of the specimen.
Figure 1. External anatomy of a generalized caterpillar. Redrawn and modified from Snodgrass (1935). Lepid63L.gif
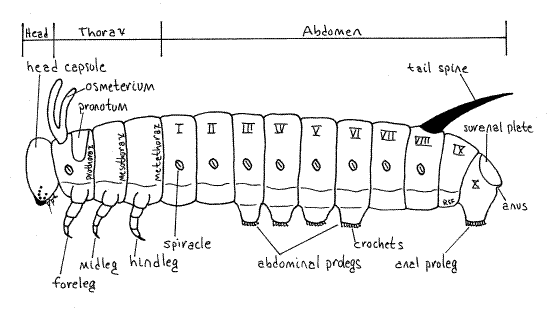
The larva is, naturally, eruciform, or caterpillar-like having a more or less cylindrical body and well-developed sclerotized head capsule, thoracic legs and abdominal prolegs (Fig. 1). The body is smooth and naked, with inconspicuous, scattered, short setae, visible only with magnification. The body, like that of other insects, consists of a small anterior head, middle thorax, and large posterior abdomen. Together the thorax and abdomen are the trunk. The head capsule is wider than the short neck, or cervix, connecting it with the thorax, a characteristic of the family Hesperiidae. Paired, jointed, segmental appendages are present on the head and thorax but are lacking on the caterpillar abdomen. Paired, unjointed, segmental, fleshy prolegs, however, are found on the abdomen. The body wall is transparent and the insect is the color of the vegetation in the gut.
A healthy specimen in a dish with a piece of fresh canna leaf will probably construct a domicile, or leaf nest. Watch as the larva touches its “chin” (with the spinneret) to the leaf surface to attach a silken thread on one side of the developing nest. It then swings the head to the other side of the leaf and again touches the spinneret to the leaf to anchor the other end of the thread. The silk shrinks and hardens upon exposure to air, drawing the edges of the nest together to form a tube enclosing the animal.
Head
The head is enclosed in a rigid sclerotized head capsule, or epicranium, which may be black or orange, depending on instar. The head consists of several fused segments and bears the appendages of those segments. The component segments of the head are not recognizable and it appears to be unsegmented.
Dorsally the epicranium is divided into large right and left halves by a Y-shaped line, the epicranial suture. The forks of the “Y” are made of the two frontal sutures whereas the stem is the coronal suture (Fig. 2). The lateral regions of the epicranium are the cheeks, or genae, whereas the dorsal posterior portion is the vertex. The triangular frons and the clypeus lie between the two frontal sutures. The frons is dorsal and clypeus is situated on its ventral border. In fifth instar larvae the epicranium is orange or beige and the frons bears a black spot. The bilobed labrum articulates with the ventral edge of the clypeus. The labrum is the anterior border of the preoral cavity, which is a space surrounding the mouth and formed by the mouthparts; labrum anteriorly, mandibles and maxillae laterally, and labium posteriorly (Fig 21-7*).
Figure 2. Dorsal view of the head capsule of the Greater Canna Leafroller, Calpodes ethlius. Lepif54L.gif
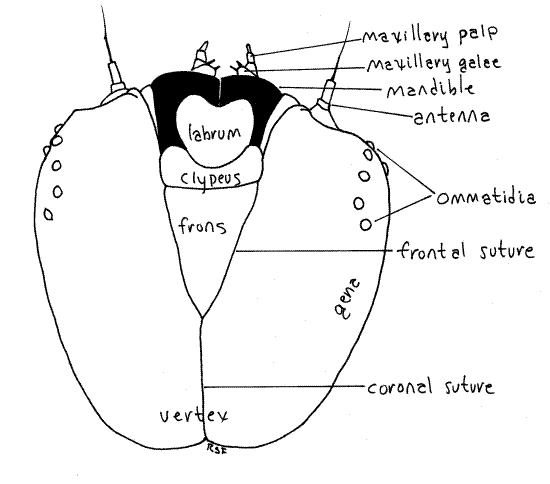
Posterior and ventral to the labrum is a pair of large, dark mandibles each with a smooth, curved, toothless blade resembling a sculptor’s or woodworker’s gouge. The blades of the two mandibles oppose each other on the midline and are used to cut discs of tissue from the canna leaf. If your specimen is alive and active you will probably see the mandibles moving in the transverse plane. The mandibles form the sides of the preoral cavity and lie beside the mouth.
The maxillae and labium are small, inconspicuous, and best studied after the specimen is anesthetized. Immediately posterior to each mandible is a maxilla from which protrudes a small galea and a slightly larger maxillary palp (Fig. 2-4). The galea is tipped with several setae whereas the palp is biarticulate. During metamorphosis the galea becomes the large sucking proboscis characteristic of adult butterflies and moths. The labium lies on the midline between the two maxillae and bears the median spinneret. This is a hollow spine at whose tip the salivary (silk) duct from the salivary (silk) glands opens. A tiny, vestigial labial palp protrudes on each side of the spinneret. The labial palps become greatly enlarged in adults and, along with the maxillary proboscis, are the only functional adult mouthparts.
A pair of short, inconspicuous, pale, cylindrical antennae can be seen lateral to the mandibles, between the mandibles and ommatidia (Fig. 2, 3). The antennae and ommatidia should be revisited after you have anesthetized the specimen and it ceases to move.
Figure 3. Ventral view of the head capsule of the Greater Canna Leafroller, Calpodes ethlius. Lepid55L.gif
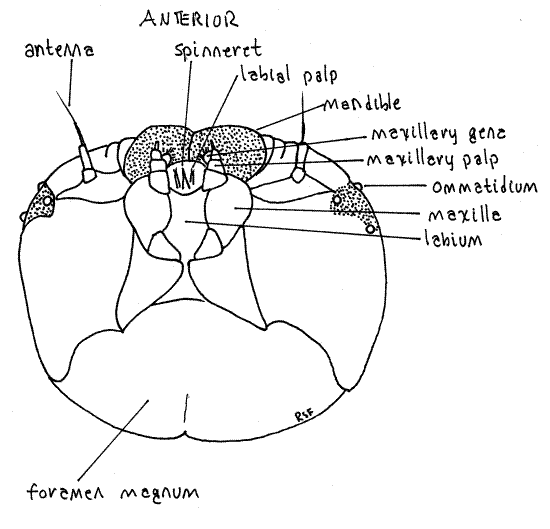
Posteriorly the head capsule is penetrated by a large opening, the foramen magnum, through which the gut, nerve cord, silk ducts, aorta, hemocoel, and tracheae pass between head and thorax (Fig. 3). The foramen is filled with soft tissue and is cannot be seen when the head is attached to the thorax but you can make out its outline by pushing the soft tissue of the cervix aside.
Figure 4. Lateral view of the right side of the head capsule of the Greater Canna Leafroller, Calpodes ethlius. Lepid56L.gif
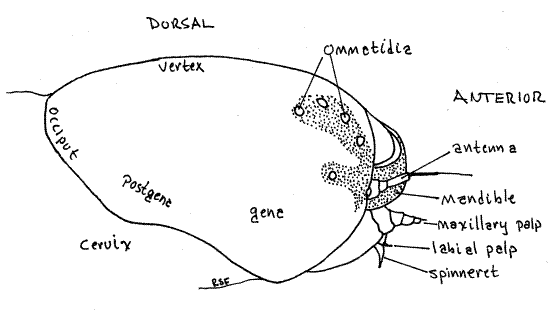
Antero-laterally each side of the epicranium bears a pigmented area with the colorless, transparent corneae, or lenses, of six large, non-contiguous ommatidia, sometimes erroneously referred to as ocelli but there are no true ocelli in caterpillars (or adult butterflies) and the ommatidia are derived from a compound eye. Together they are known as the ocularium.
Thorax
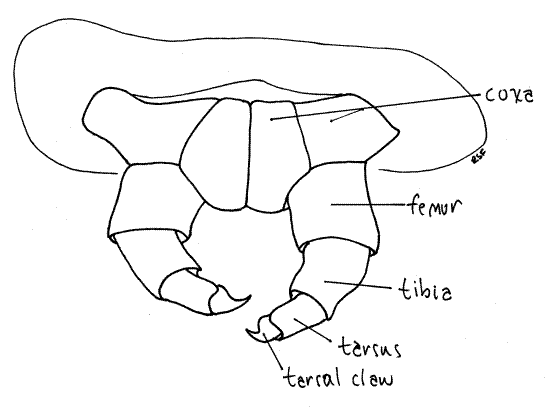
The insect thorax consists of three segments, prothorax, mesothorax, and metathorax, each of which bears a pair of sclerotized, jointed thoracic legs (Fig. 1, 16-2). These are theforelegs, midlegs, and hindlegs, respectively. Each thoracic leg consists of a series of cuticular rings decreasing in size distally. The largest ring is the proximal coxa, which articulates with the body Fig. 5, 21-1E). It is followed in turn by the femur, which is fused with a vestigial trochanter, then the tibia, and finally a biarticulate tarsus. The terminal tarsal article is a darkened, sclerotized claw.
The thorax is connected with the head by a short unsclerotized cervix, or neck, which in hesperiids is not as wide as either the head or prothorax. The thoracic segments increase progressively in width posteriorly with the prothorax being about the width of the head. The prothorax bears a cuticularized pronotum, or prothoracic shield, covering its dorsal and lateral surfaces. A dark brown or black spot marks each lateral edge of the pronotum. A white prothoracic spiracle can be seen on each side near the edge of the shield (Fig. 1). These are actually the spiracles of the mesothoracic segment that have migrated anteriorly. The mesothorax and metathorax have neither sclerotized nota nor spiracles.
Figure 5. Forelegs of the Catalpa Caterpillar, Ceratomia catalpae. Lepid59L.gif
Abdomen
The abdomen (Fig. 1) is a succession of ten segments which increase posteriorly in diameter to segments five and six, after which they decrease. The result is a caterpillar tapered at both ends. Most of the abdomen is loaf-shaped (round above, flat ventrally) in cross section but posteriorly it is flattened. Abdominal segments 3-6 and 10 each bear a pair of soft abdominalprolegs (Fig. 1). Those of the 10 th segment are the anal prolegs. The prolegs make good landmarks for recognizing the segments by number. Each proleg terminates in a central sucker surrounded by a circle of sclerotized hooks, or crochets. The suckers can be used to grasp smooth surfaces whereas the crochets function chiefly in attaching to silk or other rough surfaces. Under the influence of normal blood pressures the crochets engage the substratum and attach. Elevated blood pressure causes the crochets to disengage. Consequently, at rest the larva is by default attached to is substratum and unlikely to fall. An active elevation of hemocoelic pressure is required to disengage.
Abdominal segments 1-8 each bear a pair of oval white spiracles which open into the common tracheal system (Fig. 1). A typical insect respiratory system of interconnected tracheae is clearly visible through the body wall as a series of white tubules in the body cavity (hemocoel) between the body wall and the gut. Tracheae arise from each spiracle and branch dendritically (tree-like) to supply nearby tissues and organs with oxygen (Fig. 6). Most conspicuous are the three sunbursts of branching tracheae radiating outward from each spiracle and the two unbranched lateral longitudinal tracheal trunks, which connect successive spiracles on each side of the body.
The tracheal system is complex and ramifies throughout the body to provide oxygen and remove carbon dioxide to and from all tissues. The hemal system plays no role in oxygen transport in insects. Pay attention as you dissect the caterpillar to the pervasiveness of the tracheal system. You will find it extending to all tissues.
Figure 6. Cross section of a typical insect abdominal segment. Redrawn from Snodgrass (1935). Lepid65L.gif
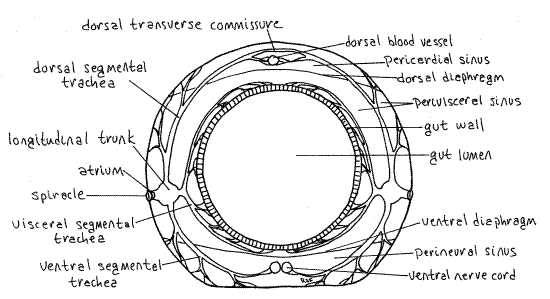
In Calpodes many aspects of the tracheal system are visible through the body wall and dissection is not required to see it. In general, each spiracle opens into a short atrium that connects with the lateral longitudinal tracheal trunk (Fig. 6). From this junction arise three major segmental tracheae, each with many branches. Together these are the three sunbursts mentioned above. The dorsal segmental trachea and its many branches extend to the dorsal body wall and heart musculature. The visceral segmental trachea sends branches to the gut, fat body, silk glands, and gonads. The ventral segmental trachea supplies the ventral body wall musculature and nerve cord with oxygen. As you observe an active caterpillar from the side, the dorsal and ventral branches remain stationary as the gut moves but the visceral branch moves with the gut.
In some segments the right and left dorsal segmental tracheae join each other across the dorsal midline to form a dorsal transverse tracheal commissure. Such a dorsal transverse commissure can be seen connecting the spiracles of the eighth abdominal segment. Similarly some ventral segmental tracheae form ventral transverse commissures. There may be additional longitudinal trunks associated with the gut and dorsal body wall. These are smaller than the lateral longitudinal tracheal trunks. Find as many of these branches as possible. You will have another opportunity to study the tracheal system after you open the hemocoel but most of its major features can be seen now, prior to dissection.
Note that, because of the numerous longitudinal and transverse connections, the tracheae form a single interconnected system so that all parts of the body, whether or not they have their own spiracles, are supplied with tracheae and oxygen. The head, two thoracic, and two abdominal segments, for example, lack spiracles but nevertheless are provided with oxygen by the system.
The gut through most of its length is only slightly smaller in diameter than the caterpillar itself and can be clearly seen in constant motion just inside the body wall. If the caterpillar has fed recently, the gut will be filled with pieces of vegetation of uniform (bite) size. These leaf fragments are cut from the host plant by the curved blades of the mandibles. Posteriorly these are formed into larger, darker, loosely consolidated fecal pellets consisting of these same pieces of vegetation compacted into pellets and packages in a membrane. There is no triturative proventriculus in Calpodes and consequently the bites produced by the mandibles retain their original shape for the entire length of the system, from mouth to anus. The gut visible at present is mostly the crop, midgut, and hindgut but they are difficult to distinguish prior to dissection. The crop occupies most of the thorax and the midgut extends from the posterior thorax through the anterior abdomen to about the level of abdominal segment six. The hindgut is in abdominal segments 7-10 and is divided into an intestine in segments 7-8 and a rectum in segments 9-10. The rectum opens through the anus on segment 10.
The central feature of the insect hemal system is a tubular dorsal blood vessel that extends the length of the animal immediately beneath the dorsal midline (Fig 16-7). Posteriorly the dorsal vessel is contractile and is the heart whereas anteriorly it is not contractile and is known as the aorta. The heart consists of a series of sequential segmental expansions, or chambers, each with a pair of ostia. Because the body wall is transparent, the heart and aorta are easily visible in living specimens. In such a specimen you can observe the conspicuous peristaltic waves passing from posterior to anterior along the heart. The heart and aorta are enclosed in the pericardial sinus.
The insect body cavity is the hemocoel, which is filled with circulating blood that bathes the tissues and transports food, wastes, and hormones (but not oxygen). Most organ systems, including the gut, nerve cord, and excretory organs, are in the hemocoel. The hemocoel is divided by two horizontal septa (dorsal and ventral diaphragms) into three sinuses. The dorsal pericardial sinus encloses the heart, the middle perivisceral sinus is the largest and contains the gut and other viscera. The small, ventral perineural sinus surrounds the ventral nerve cord. The lateral borders of the pericardial sinus are visible through the transparent body wall on each side of the heart.
In late instar male larvae a pair of irregularly shaped, pale, yellowish testes are easily seen, even without magnification, beside the heart on the anterior border of the sixth abdominal segment.
The long, looped, slender, yellow or white Malpighian tubules arise from the midgut-hindgut junction and are visible laterally in the abdomen (Fig 16-9.
Segments 7 and 8 bear the paired pores of powder glands whose secretions form conspicuous, bright white patches of waxy powder on the ventral surfaces of these segments. This powder is used to waterproof the pupa and pupal nest and is typical of skipper larvae.
Segment 9 lacks either spiracles or prolegs and is short and inconspicuous (Fig. 1). Segment 10 is dorso-ventrally flattened, bears a pair of anal prolegs and is covered by a dorsal sclerotized, but not darkened, suranal sclerite (suranal plate). The anus lies hidden under the suranal sclerite. The anal comb is a fan-shaped, toothed sclerite dorsal to the anus, under the suranal sclerite. It can be seen by lifting the sclerite. It is used to flick fecal pellets away from the nest.
If your caterpillar is living, it may present you with one or more fecal pellets. If so, dissect one at about 20X to see that it is enclosed in a delicate peritrophic membrane. Inside the membrane you will find the uniformly sized and shaped bites of tissue cut from the host plant by the mandibles.
Internal Anatomy
High Magnification Review of External Anatomy
The remainder of the exercise should be performed on an anesthetized or recently sacrificed specimen. It makes little difference which. Anesthetize (or kill) a caterpillar by immersing it in 7% ethanol (non-denatured) in a small, wax-bottom dissecting pan. The animal will succumb in 5-10 minutes. Dissecting pans made from anchovy fillet tins are an ideal size for most insects and fit conveniently on the stage of a dissecting microscope.
Spend a few minutes observing features that were difficult to see, such as the head appendages, spiracles, thoracic legs, prolegs, and crochets earlier while the animal was active. Reexamine the heart, find the Malpighian tubules, and look once again at the details of the tracheal system.
If your specimen shows any signs of life it may periodically extend and retract the crochets of the prolegs. Watch a circle of crochets under high magnification of a dissecting microscope during one of these cycles. At rest the curved crochets are deployed and would engage silk strands or another rough substratum. When withdrawn they release their hold on the silk and permit movement of the proleg.
Examine a spiracle at about 40X. Each spiracle consists of a brownish, sclerotized, cuticular ring surrounding a recessed atrium from which a short spiracular trachea connects with the longitudinal trunk. The atrium is covered and protected by a filter apparatus consisting of a ring of closely spaced setae. The atrium is under the filter and cannot be seen from the surface. A slit-like atrial orifice penetrates the center of the filter and opens into the atrium below it. Insert a minuten nadel into the orifice to demonstrate its presence and continuity with the atrium. Use the nadel to demonstrate that the filter is composed of bristles extending into the orifice from the surrounding ring. In most insects, including Calpodes, the spiracle is equipped with a valve, under muscular control, that can close the spiracle to conserve water.
Find the articles (coxa, femur, tibia, biarticulate tarsus including terminal claw) of a thoracic leg using 40X. Examine the anal comb with 40X.
Dissection
" To begin the dissection, insert one blade of a fine scissors under the posterior edge of the suranal sclerite and cut anteriorly through the body wall well to the right of the middorsal line so that the median heart and pericardial sinus are not damaged. The heart will end up on the left side of the incision. Remember this so you can find it later after it is pinned to the bottom of the dissecting pan. Hemocoelic pressure may force a bulge of the gut through the incision but this problem will diminish as the incision is lengthened. Avoid cutting deeper than the body wall and be careful that your cut does not penetrate the gut wall. Extend the incision anteriorly to the posterior border of the head capsule but do not cut into the capsule. Use #1 stainless steel insect pins to pin the edges of the body wall against the floor of the dissecting pan. Begin at the posterior end and work your way anteriorly, inserting each pin at a 45 ° angle and stretching the body wall laterally and posteriorly as you pin it. Much of the space in the hemocoel is filled with the white ribbon-like fat body, tracheae, and gut. It may be necessary to detach some of the fat body from the body wall in order to separate it from the gut and pin it.
The body cavity opened by this incision is the perivisceral sinus, a division of the hemocoel. It is filled with blood (hemolymph) (Fig 6, 16-7). The much smaller pericardial sinus, which is also filled with blood, surrounds the heart and has been pinned aside with the body wall. Organs and tissues, including the heart, gut, fat bodies, salivary glands, excretory organs, and nerve cord, are suspended in the hemocoel and surrounded by blood.
Hemal System
The insect hemal system consists of a tubular dorsal blood vessel extending the length of the body immediately under the dorsal midline (Fig. 7, 16-7). Posteriorly, for most of the length of the abdomen, the vessel is the muscular, contractile, and ostiate heart. The vessel continues anteriorly through the thorax to the head as the non-contractile aorta.
You should have observed the heart earlier before anesthetizing and dissecting the caterpillar. Take another look at it now. It should still be present in the pericardial sinus attached to the body wall and pinned to the wax of the dissecting pan on the left side of your specimen. It may still be beating weakly. The heart is a transparent, colorless tube on the dorsal midline. It would be difficult to see were it not for the parallel rows of opaque, white, irregularly shaped nephrocytes on its walls. These special nephrocytes are large cells known as pericardial cells. The heart can be recognized as a clear channel between two parallel rows of nephrocytes. A pair of large, white, ribbon-like fat bodies lie beside the heart and may lie over it and obscure your view. That portion of the dorsal vessel in the thorax is the aorta. Remove some of the fat body if necessary to improve your view of the heart and aorta and also to reveal the dorsal body wall muscles. Large, translucent, strap-like longitudinal body wall muscles lie on either side of the heart and are reminiscent of the continuous muscle layers of an annelid rather than the individual muscles of an arthropod.
Fat Body
In Calpodes the fat bodies are conspicuous in the perivisceral sinus as longitudinal, white, convoluted ribbons attached to the body wall and gut tube. Note the tracheae extending to the fat bodies.
The fat body is a large, multipurpose, mesodermal organ consisting of various types of cells involved in intermediary metabolism, protein synthesis, and storage of lipids, carbohydrates, proteins, uric acid, and energy. In most insects the chief period of feeding, growth, and energy storage is as juveniles, for adults feed relatively little or not at all. The energy accumulated by juveniles is stored in the fat body and later used to support the activities, chiefly dispersal and reproduction, of the imago.
Fat bodies are conspicuous in the perivisceral sinus as seemingly amorphous white or yellow leafy sheets attached to the body wall and extending throughout the hemocoel. The system typically consists of a peripheral fat body forming a layer attached to the body wall and a perivisceral fat body associated with the gut wall as well as smaller concentrations with specific organs. Note the tracheae extending to the fat bodies. These thin sheets, each only one or two cells thick, expose a large surface area to the hemolymph. All cells in contact with the blood for the exchange of materials.
Digestive System
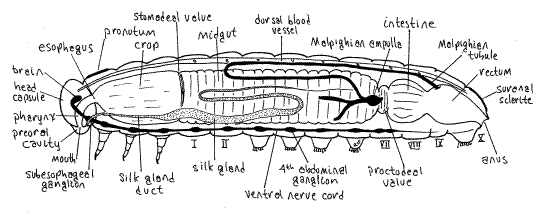
The digestive system consists of anterior ectodermal foregut, middle endodermal midgut, and posterior ectodermal hindgut, which may themselves be regionally specialized. Note the numerous branches of the visceral tracheal system extending to the gut. The foregut of Calpodes, which is lined by cuticle, consists of mouth, pharynx, esophagus, and crop. Its role is mostly in food storage with some preliminary mechanical and chemical digestion. The pharynx and esophagus are of small diameter and are largely enclosed within the head capsule. Theesophagus can be seen emerging from the capsule where it widens to become the crop at the anterior end of prothorax (Fig. 7). The crop is a storage chamber occupying most of the space in the thoracic perivisceral sinus. Its walls are transparent and its diameter is almost equal to that of the thorax. It is probably full of recently ingested food particles cut from the host plant by the mandibles.
The midgut is about the same diameter as the crop and occupies most of the anterior abdomen from segment 1 posteriorly to about segment 7. Unlike that of the crop, its wall is opaque. The chief functions of the midgut are enzyme secretion, hydrolysis, and absorption.
The midgut narrows abruptly in segment 7 to form the proctodeal valve (pyloric valve) which separates midgut from hindgut. The hindgut, which is cuticularized like the foregut, consists of a short intestine (anterior intestine), a short rectum (posterior intestine), and the anus. The intestine and rectum are separated from each other by a constriction. The hindgut functions in water and salts reclamation, a vital role in terrestrial animals, and feces formation and storage. Later you will open the gut tube to study its interior.
Figure 7. Internal anatomy of a generalized caterpillar viewed from the left. Two of the three left side Malpighian tubules have been truncated for clarity. Redrawn and modified from Snodgrass (1935). Lepid64L.gif
A pair of glands opening on the labium and thus known as the labial glands function in most insects as salivary glands and secrete saliva with hydrolytic enzymes adapted for the diet of the species. Such labial glands are sometimes referred to as salivary glands. In some taxa, most notably Lepidoptera (butterflies and moths), Trichoptera (caddisflies), and Hymenoptera (bees, ants, and wasps), the larval labial glands secrete, not saliva, but a proteinaceous silk used for a variety of purposes and are known as silk glands. In addition, caterpillars have a pair of mandibular glands, opening on the mandibular segment, that secrete saliva whose enzymes assist in the digestive process. The mandibular glands probably will not be seen.
In Calpodes each silk gland is a long, transparent or opaque, colorless, white, or yellowish-brown tube lying to the right or left of the gut in the ventral perivisceral hemocoel (Fig. 7). A common duct from the spinneret arises within the head capsule, where you cannot see it, bifurcates to form the right and left silk gland ducts. These narrow, transparent ducts can be seen emerging laterally from the foramen magnum to enter the thorax. The ducts are small in diameter but quickly expand to become the much larger silk glands. Each gland extends to the posterior abdomen, then reverses direction and extends back to the anterior abdomen, only to reverse direction once more and run posteriorly again, now closely associated with the fat body and smaller in diameter. The opaque central region of the gland is the largest in diameter and is well supplied with tracheae. In the caterpillar of the silk moth, Bombyx mori, secretory cells in the epithelium of the posterior region of the gland secrete fibroin, the major silk protein, composed of about 40% glycine. The central region of the gland secretes other proteins whose function is to coat the fibroin strands and hold them together.
Respiratory System
The tracheal system was studied prior to dissection (Fig 6). You may now review its major features and look for some of the details you could not find earlier. Study the internal aspect of a spiracle with 40X and find the major tracheal branches.
" Use fine scissors to remove a piece of a longitudinal trunk and make a wet mount with it. Examine the preparation with 100X, then 400X of the compound microscope. Note the chitinous rings, known as taenidia, that reinforce the walls of the trachea and hold it open, much like the cartilaginous rings that hold your own trachea open (Fig 21-10A).
Excretory System
The excretory system consists of nephrocytes in the hemocoel and Malpighian tubules, also in the hemocoel. Lepidopteran larvae and adults have six Malpighian tubules, three arising on each side, right and left, of the midgut-hindgut junction in the vicinity of the proctodeal valve at the constriction between midgut and intestine (Fig. 7). On each side the cluster of three arises from a common ampulla evaginated from the gut. The tubules are supplied with tracheae. It is difficult to demonstrate the connection of the Malpighian tubules with the gut in gross dissection.
The tubules emerge from the gut at about the level of abdominal segment 6 and extend anteriorly as long, slender, opaque white or yellow tubules to the anterior abdomen. After reaching their anteriormost position they reverse direction and loop posteriorly, sometimes as far as the posterior end of the abdomen. To further complicate the pattern in Lepidoptera, the distal ends of the six tubules enter the connective tissue of the rectum wall and end there, out of sight, and there are thus no free ends of the tubules. The connections of both ends of the Malpighian tubules with the gut are difficult to demonstrate in gross dissection.
Nephrocytes are scattered throughout the hemocoel and are usually not visible in gross dissection. Nephrocytes are thought to be storage kidneys that absorb and sequester a variety of particles and dyes but do not phagocytose bacteria. Special large nephrocytes, known as pericardial cells, form a row on each side of the heart. You saw them earlier.
Digestive System Internal Anatomy
" Make a longitudinal, middorsal incision along the entire length of the exposed gut to open its lumen, which is probably filled with small pieces of canna leaf. In the midgut the food mass of leaf particles is enclosed in a thin, transparent peritrophic membrane (Fig 21-9). Cut through the peritrophic membrane and use a Pasteur pipet to remove the leaf fragments and improve your view of the gut lumen. The foregut (crop) is separated from the midgut by an inconspicuous stomodeal valve (cardiac valve), which in Calpodes, is manifest as a slight brownish thickening of the gut wall (Fig. 7). In many insects the valve is more elaborate. The proctodeal valve, between the midgut and hindgut, is a more substantial thickening of the gut wall. Find the anus from inside the rectum and note the appearance of the anal comb from this viewpoint.
The peritrophic membrane is secreted by cells in the vicinity of the stomodeal valve and thus there is no peritrophic membrane anterior to the foregut-midgut junction. The peritrophic membrane is secreted by the epithelium of the anterior midgut epithelium to enclose the food mass.
The opacity of the midgut wall is due to its mucosal epithelium. The epithelium has a brush border of microvilli that extend through the peritrophic membrane into the gut lumen to absorb food molecules. This border is visible in fresh specimens as a bluish green iridescence.
Nervous System
In its primitive, largely uncephalized condition, the central nervous system includes the tripartite brain, or supraesophageal ganglion, consisting of proto-, deuto- and tritocerebrum and the ventral nerve cord consisting of segmental ganglia joined by paired longitudinal connectives (Fig 16-11). The brain innervates the eyes (protocerebrum), antennae (deutocerebrum), and labrum (tritocerebrum). The tritocerebrum is connected with the double longitudinal ventral nerve cord by a pair of circumesophageal connectives. Immediately posterior to the connectives is the subesophageal ganglion consisting of the fused ganglia of the mandibles, maxillae, and labia. The segmental ganglia of the thorax and abdomen remain independent of each other and are spaced along the nerve cord. The CNS of Calpodes exhibits this primitive condition in most respects and shows little cephalization or condensation of ganglia.
" Cut the tracheae on the right side of the gut, move the gut to the left, and pin it so the ventral midline is revealed. Remove the large, lateral fat bodies as necessary to reveal the ventral midline of the thorax and abdomen, being careful that you do not damage the nerve cord on that midline. Do not attempt to remove the small ventral fat bodies associated with the nerve cord.
The segmental ganglia are bright white and easy to see widely spaced along the ventral nerve cord. The cord itself is not as obvious as the ganglia but once you find the ganglia you can find the nerve cord connecting them. In the abdomen the parallel connectives are fused and the double nature of the nerve cord is not apparent although in the thorax the connectives are widely separated and the cord is quite clearly double.
The subesophageal and first thoracic ganglia are far anterior and can best be seen by lifting the head capsule and looking into the foramen magnum using 20X. The subesophageal ganglion, which serves the mandibles, maxillae, and labium, is in the posterior head capsule but it can be seen lying below the gut by looking into the foramen magnum (Fig. 7, 21-7). The brainmay also be visible, but dorsal to the gut. The subesophageal ganglion is connected by a pair of short connectives with the first thoracic ganglion, which is very close to it. The second thoracic ganglion is well separated from the first and connected to it by a pair of long connectives. The third thoracic ganglion is far posterior to the second and is connected with it by a pair of long connectives.
The first abdominal ganglion is very close to the third thoracic so the connectives between them are very short. Abdominal ganglia 2-7 are widely spaced along the length of the abdomen and are connected by coalesced connectives, which form the double nerve cord, which appears to be single (Fig. 6). The abdominal part of the nerve cord is partly hidden by the ventral fat bodies. Abdominal ganglion eight is very close to the seventh ganglion and their connectives are very short. Several nerves radiate posteriorly from the eighth ganglion. The combined connectives, whether single or double, form the ventral nerve cord.
Examine a ganglion, such as the first abdominal, with 40X and find the pair of inconspicuous segmental nerves connecting the ganglion with the periphery. Note that the nerve cord, ganglia, and nerves are supplied with tracheae.
Note that ventral longitudinal body wall muscles, similar to the dorsal muscles seen beside the heart, lie beside the nerve cord.
Reproductive System
The reproductive system of caterpillars is rudimentary and will develop in the pupa during metamorphosis. Small gonads, however, are present in late instar larvae and spermatogenesis occurs in these larvae and in the pupa. Eggs develop in the pupa. Development of both internal and external genitalia occurs during metamorphosis in the pupa.
*Hyphenated figure call-outs, such as this one, refer to figures in Ruppert, Fox, and Barnes (2004). Those without hyphenation refer to figures embedded in this exercise.
References
Borror DJ, Triplehorn CA, Johnson NF . An introduction to the study of insects, 6 th ed. Saunders College Publishing, Philadelphia. 875pp.
Chapman RF. 1998. The insects, Structure and function, 4 th ed. Cambridge Univ. Press, Cambridge. 769 pp.
Comstock JH. 1930. An introduction to entomology. Comstock Pub., Ithaca. 1044 pp.
McAuslane HJ, King K. 2000. Larger Canna Leafroller. Integrated Pest Management, Featured Creatures. University of Florida, Department of Entomology and Nematology. Institute of Food and Agricultural Sciences.http://ipm.ifas.ufl.edu/ctgysrch/insects.htm#fc
King HS. 1880. Internal organization of Hesperia ethlius Cram. As observed in the living animal. Psyche 3:322-324.
Ross HH. 1965. A textbook of entomology, 3 rd ed. John Wiley & Sons, New York. 539pp.
Ruppert EE, Fox RS, Barnes RB. 2004. Invertebrate Zoology, A functional evolutionary approach, 7 th ed. Brooks Cole Thomson, Belmont CA. 963 pp.
Scott JA. 1986. The butterflies of North America. Stanford University Press, Stanford. 583pp.
Snodgrass RE . 1935. Principles of insect morphology. McGraw-Hill, New York. 667 pp.
Supplies
1 1iving, freshly collected, late instar Calpodes ethlius caterpillar
1 small (anchovy tin) wax-bottom dissecting pan
1 dissecting microscope
1 compound microscope (can be shared by several students or the entire class)
1 microscope slide and coverslip
1 microdissecting forceps
1 microdissecting (iridectomy) scissors
2 minuten nadeln with applicator stick handles
20 # 1 stainless steel insect pins
7% non-denatured ethyl alcohol
1 plastic Pasteur pipet