Invertebrate Anatomy OnLine
Ammotrechella stimpsoni ©
Camel Spider
20jun2006
Copyright 2001 by
Richard Fox
Lander University
Preface
This is one of many exercises available from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises can be accessed by clicking on the links to the left. A glossary and chapters on supplies and laboratory techniques are also available. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Arthropoda P, Chelicerata sP, Euchelicerata, Arachnida C, Solifugae O, Ammotrechidae F (Fig 16-15, 18-47)
Arthropoda P
Arthropoda, by far the largest and most diverse animal taxon, includes chelicerates, insects, myriapods, and crustaceans as well as many extinct taxa such as Trilobitomorpha. The segmented body primitively bears a pair of jointed appendages on each segment. The epidermis secretes a complex cuticular exoskeleton which must be molted to permit increase in size. Extant arthropods exhibit regional specialization in the structure and function of segments and appendages but the ancestor probably had similar appendages on all segments. The body is typically divided into a head and trunk, of which the trunk is often further divided into thorax and abdomen.
The gut consists of foregut, midgut, and hindgut and extends the length of the body from anterior mouth to posterior anus. Foregut and hindgut are epidermal invaginations, being derived from the embryonic stomodeum and proctodeum respectively, and are lined by cuticle, as are all epidermal surfaces of arthropods. The midgut is endodermal and is responsible for most enzyme secretion, hydrolysis, and absorption.
The coelom is reduced to small spaces associated with the gonads and kidney. The functional body cavity is a spacious hemocoel divided by a horizontal diaphragm into a dorsal pericardial sinus and a much larger perivisceral sinus. Sometimes there is a small ventral perineural sinus surrounding the ventral nerve cord.
The hemal system includes a dorsal, contractile, tubular, ostiate heart that pumps blood to the hemocoel. Excretory organs vary with taxon and include Malpighian tubules, saccate nephridia, and nephrocytes. Respiratory organs also vary with taxon and include many types of gills, book lungs, and tracheae.
The nervous system consists of a dorsal, anterior brain of two or three pairs of ganglia, circumenteric connectives, and a paired ventral nerve cord with segmental ganglia and segmental peripheral nerves. Various degrees of condensation and cephalization are found in different taxa.
Development is derived with centrolecithal eggs and superficial cleavage. There is frequently a larva although development is direct in many. Juveniles pass through a series of instars separated by molts until reaching the adult size and reproductive condition. At this time molting and growth may cease or continue, depending on taxon.
Chelicerata sP
Chelicerata is a large taxon that includes spiders, scorpions, pseudoscorpions, ticks, mites, horseshoe crabs, sea spiders, and many others. The group originated in marine habitats but almost all modern chelicerates are terrestrial.
The body is divided into an anterior cephalothorax with six pairs of appendages and a posterior abdomen which, in most groups, does not bear appendages or has highly modified appendages. The first appendages of the cephalothorax are the chelicerae. Antennae are not present and the brain has two regions rather than the three found in mandibulates. Appendages are primitively biramous but are uniramous in almost all Recent taxa.
Euchelicerata
The segments of the cephalothorax are fused and covered by a dorsal shield, or carapace. Two median eyes are present.
Arachnida C
Arachnids are the terrestrial descendents of the early aquatic chelicerates. The taxon includes the mites, scorpions, pseudoscorpions, spiders, harvestmen, and several other taxa. All are adapted for a terrestrial existence with internal gas exchange and a waterproof integument. Some are capable of silk production and many use toxins to subdue or kill the prey. Most are carnivores, digestion is usually outside the body, and food is liquefied before ingestion.
Solifugae O
The solifuges (or solpugids) are the camel spiders (also known as wind scorpions or sun spiders showing a curious mixture of primitive and derived arachnid characters. Most are fast-moving, nocturnal raptors with a superficial resemblance to spiders. The size ranges from a few millimeters to giants up to 7 cm long. The order comprises 12 families, 153 genera and 981 described extant species. Only two families, Ammotrechidae and Eremobatidae, occur in the western hemisphere and only Ammotrechidae is represented in eastern North America. About 120 occur in North America, mostly in the arid west. Ammotrechella stimpsoni, which occurs in Florida and the West Indies, is the only eastern North American species. It is nocturnal and feeds termites (Reticulitermes flaviceps), as do most American species. Most solifuges eat insects but some are known to consume lizards. Solifuges have very large chelicerae and long, slender, leglike pedipalps. Most are very setose. The anterior cephalothorax is covered dorsally by a carapace but its posterior segments are free of the carapace and covered by individual tergites. The biarticulate chelicerae are gigantic in comparison with the body. The pedipalps are long and leglike, with an eversible adhesive tip. The first walking legs are antenniform with a sensory role, rather than locomotory, role. The abdomen is segmented with segmental tergites and sternites. There is no telson and no pedicel. They differ from spiders, which they resemble, in their partially segmented cephalothorax, segmented abdomen, and the absence of venom, silk glands or spinnerets. They are sometimes known as “false spiders” because of this resemblance. This exercise is based on a single specimen captured in Ft. Pierce, Florida in July 1990.
External Anatomy
Tagmata
Cephalothorax
The solifuge body consists of an anterior cephalothorax (= prosoma) and posterior abdomen (= opisthoma) demarcated (Fig 1, 18-36). Both tagmata are segmented.
The cephalothorax is composed of six segments numbered 1-6 from anterior to posterior (plus a vestigial seventh). The anterior region consisting of the first four segments, is covered by a large tergite, the anterior carapace (= propeltidium). The anterior carapace is strongly humped dorsally, a feature responsible for the name “camel-spider”. The two posterior cephalothoracic tergites (5 and 6) are free of the anterior carapace and are covered by separate tergites (mesopeltidium, metapeltidium respectively). The segmentation of the anterior cephalothorax is not evident dorsally. Segments 5 and 6, however, are distinct from each other and from the anterior cephalothorax. They resemble the abdominal segments but differ from them in possessing appendages.
The anterior carapace bears a pair of large median eyes on its anterior margin (Fig 1, 18-36B). Vestigial eyes may be present on the sides of the carapace.
The cephalothorax bears the six pairs of appendages characteristic of chelicerates. The first appendages are the large biarticulate chelicerae. Together they form a large cone that projects anteriorly from the front of the head. In relation to the side of the body, they are larger than those of any other arachnid.
Each chelicera is formed of two articles which together form a powerful pincer whose jaws open and close in the vertical plane. The two chelicerae are held close together and their medial surfaces are flattened accordingly. A hissing noise is made by rubbing the chelicerae together. Solifuges can bite but they have no poison glands. Solifuges are said to have the most powerful jaws in the animal kingdom in relation to the size of the body. Males or all taxa, except Eremobatidae, have a spoon-shaped flagellum on the basal article of the chelicera. Its function is not known.
In use, the two chelicerae move forward and backward past each other. Simultaneously with this action, the fingers of each chela are also moving vertically. Prey items, such as insects, frequently termites, are cut into tiny pieces and rubbed between the distal ends of the chelicerae. They are thus quickly crushed, and liquefied into a wet shapeless mass. The muscular pharynx sucks the liquid from the mass until it is drained of its fluid and then discarded, nearly dry. The fluid and some of the tiny hard pieces are ingested.
Figure 1. Dorsal view of the solifuge, Galeodes arabs (Galeodidae). Setae have been omitted for clarity. Redrawn from Savory (1977). Solpugid9L.gif
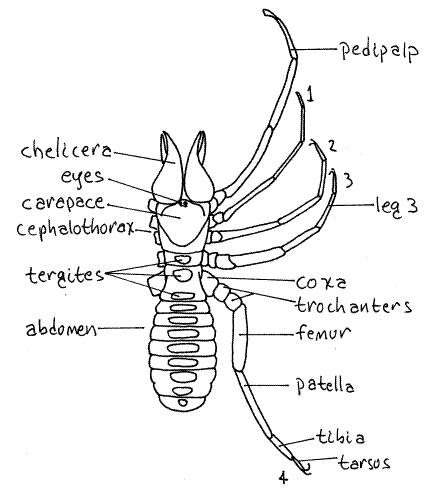
The second appendages are the pedipalps. These are very long, with six articles, most of which are slender and elongated (Fig 1). They look like legs. They are held in front of the animal and function as sense organs similar to the antennae of other arthropods. They bear chemo- and mechano-receptors and are used to locate prey. Each has an eversible adhesive disk at its blunt tip. The disks are not apparent unless they are everted. Some solifuges can climb a glass surface using only the adhesive pedipalps.
Figure 2. Ventral view of the solifuge, Rhagodes (Rhagodidae). Setae have been omitted for clarity. Redrawn from Warburton (1909). Solpugid12L.gif
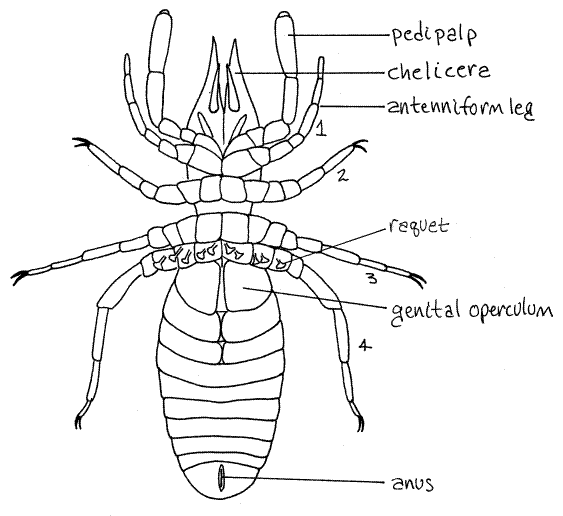
The remaining four pairs of appendages are legs composed of six or seven articles each. The articles are, in order from proximal to distal, the coxa, trochanter, femur, patella, tibia,and tarsus. The trochanters in the last three pairs of legs are divided into two parts each. The tarsi may also be divided. The number of tarsal subdivisions varies with species. Legs 2-4 bear one or two claws at the end of the tarsus.
The first pair of legs is much smaller and more slender than the others and is used as a second pair of antennae (Fig 1). It is sensory. Species of Ammotrechidae do not have claws on this leg but eremobatids do. This leg is not used for locomotion.
The remaining legs become progressively larger posteriorly and are locomotory. Male wind scorpions are very fast runners, hence the name.
Each of the posterior pair of legs is equipped with five T-shaped sensory racquets, or malleoli, on their proximal articles (Fig 2, 18-36A). These are small plates attached by short stalks to the underside of the appendage.
Abdomen
The abdomen does not bear appendages. It is soft and elongate and composed of ten free segments. Each segment has a large dorsal tergite (Fig 1) and a ventral sternite (Fig 2).
The gonopore is located midventrally on the first abdominal segment (Fig 2). It is covered by a pair of opercula thought to be derived from the appendages of this segment. The remaining abdominal segments lack appendages. The anus is ventral at the posterior end on segment 10 (Fig 2).
Respiration
Respiration, in some species, is via trachea whose spiracles open posterior to the coxae of the second legs and at the posterior edges of the sternites of abdominal segments 2 and 3 (and sometimes 4). The spiracles have opercula. Those of segments 2 and 3 are paired and located a little to the side of the ventral midline. The spiracle of segment 4 is unpaired and median if it is present.
Two large tracheal trunks extend parallel to each other just beneath the integument along the dorsal midline of the abdomen. In some species, including Ammotrechella, they can be seen through the integument of the posterior abdomen as branching white tubes. These trachea are moved toward and away from each other in periodic bursts of rhythmic pulsations that can be observed through the integument of living Ammotrechella.
The body and appendages are covered with fine long trichobothria that are sensitive to mechanical disturbances such as air movements, vibrations, and touch.
Excretion
Excretion is accomplished by Malpighian tubules and a pair of coxal glands (= saccate nephridia). The coxal glands open via nephridiopores at the base of the pedipalps.
Hemal System
The heart is a short tube in the abdomen and posterior cephalothorax. It has eight ostia (two cephalothoracic and six abdominal) but is not visible without dissection.
Nervous System
The nervous system is highly cephalized and consists of a dorsal brain and ventral subesophageal ganglion. Most of segmental ganglia are coalesced to form this large ganglion. The brain and ganglion are connected by a pair of connectives to form a circumesophageal nerve ring. A small abdominal ganglion is present.
Reproduction
The gonopore is a small slit on the sternite of the second abdominal segment. It is covered by the genital opercula (Fig 2). There is little sexual dimorphism except for the flagellum of the male chelicera.
References
Kaestner A. 1967. Invertebrate Zoology, Vol. II. Arthropod relatives, Chelicerates, Myriapoda . Wiley, New York. 472 pp.
Levi HW, Levi LR. 2002. Spiders and their kin. St. Martin’s Press, New York. 160pp.
Muma MH. 1982. Solpugida in Parker P.(ed). Synopsis and classification of living organisms . McGraw-Hill, New York. pp. 102-104.
Punzo F. 1998. The biology of camel-spiders (Arachnida: Solifugae). Kluwer Academic Pub., Boston. 301 pp.
Savory TH. 1977. Arachnida 2 nd ed. Academic Press, New York. 340p.
Ruppert EE, Fox RS, Barnes RB. 2004. Invertebrate Zoology, A functional evolutionary approach, 7 th ed. Brooks Cole Thomson, Belmont CA. 963 pp.
Vanuytven H. Arachnology Home Page : www.arachnology.be/Arachnology.html
Warburton C. 1909. Arachnida, pp. 255-473 in Harmer SF, Shipley AE (eds), Cambridge Natural History vol IV Crustacea and Arachnids. MacMillan, London.
Supplies
Living or preserved solifuge
Small dissecting pan or culture dish
Dissecting microscope