Invertebrate Anatomy OnLine
Lamprey Eel
Ammocoetes Larva ©
5jul2006
Copyright 1985 by
Richard Fox
Lander University
Preface
This is one of many exercises available from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. Additional exercises, a glossary, and chapters on supplies and laboratory techniques are also available at this site. Terminology and phylogeny used in these exercises correspond to usage in the Invertebrate Zoology textbook by Ruppert, Fox, and Barnes (2004). Hyphenated figure callouts refer to figures in the textbook. Callouts that are not hyphenated refer to figures embedded in the exercise. The glossary includes terms from this textbook as well as the laboratory exercises.
Systematics
Chordata P, Metameria, Vertebrata sP, Agnatha C, Petromyzontida O, Petromyzontidae F
Lamprey eels belong to Agnatha, the oldest known taxon of living vertebrates. Of a handful of living lampreys, the parasitic marine lamprey, Petromyzon marinus , is the most common and most readily available for laboratory use, although it is not the best example of its taxon. Agnaths are vertebrates with no paired appendages, no jaws, no cranium and no vertebrae. The skeleton is cartilaginous and the notochord persists through life. The larva is the ammocoetes, which closely resembles cephalochordates such as Branchiostoma (amphioxus). The ammocoetes (AM oh sete ease) is the vertebrate most like the most vertebrate-like invertebrate. As a vertebrate with striking similarities to a non-vertebrate it is an important evolutionary link supporting the hypothesized close relationship between protochordates and vertebrates.
Laboratory Specimens
Ammocoetes larvae of marine lampreys or brook lampreys are available from biological supply companies as wholemount slides of small specimens and cross section slides. Preserved larger larvae are also available from some suppliers. This laboratory exercise utilizes preserved specimens, wholemount and pharyngeal cross section slides and can be used with either marine or brook lamprey larvae. The larvae on wholemount slides are necessarily very small, usually less than 2 cm. Preserved specimens for this exercise should be much larger, about 100-120 mm.
The name “ammocoetes" (or ammocoete), is an old generic name retained from earlier days before we realized they were larval agnaths and were thought to be a genus of cephalochordates. The larvae live in freshwater streams, burrow in sediments, and are filter feeders utilizing phytoplankton. The larvae live two to six years before metamorphosing into parasitic adults. See Hyman (1942), Weichert (1961), or Lehman (1983) for additional information. The biology student’s first exposure to the lamprey and its ammocoetes is usually in a course in comparative vertebrate anatomy.
Anatomy
Wholemount Slide
Examine your specimen with the low power scanning lens (4X) of your compound microscope. These specimens have been cleared and stained to improve visualization of internal organs that might otherwise be visible. Note the strong resemblance the ammocoetes larva bears to amphioxus (Fig 1). Like amphioxus, it is fishlike, laterally compressed, and fusiform. Anteriorly there is a poorly defined head with an oral hood enclosing a preoral space known as the vestibule. There is a long dorsal fin and the tail bears a caudal fin.
Figure 1. Ammocoetes larva. Wholemount of a cleared specimen at 50 X. fish161L.gif
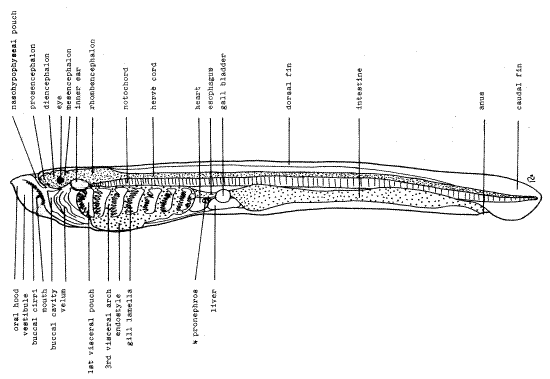
The nerve cord is a conspicuous, longitudinal, pink (stained) tube lying dorsally in the animal. Anteriorly the cord is expanded to form a distinct brain, a feature absent in amphioxus. The brain of the larva is divided into three regions, each associated with a primary sense capsule. The anterior prosencephalon is associated with the olfactory sense, the middlemesencephalon with vision, and the posterior rhombencephalon with detection of balance, acceleration, and equilibrium. The prosencephalon comprises an anterior telencephalon and a posterior diencephalon separated by a deep groove. The brain narrows abruptly posterior to the rhombencephalon to become the nerve cord. Use the 10X lens to examine the area just anterior to the prosencephalon. Here you will find a median funnel-shaped invagination of ectoderm, the nasohypophyseal pouch. It will contribute to the formation of the pituitary gland of the adult lamprey and is homologous to Rathke’s pouch and perhaps to Hatschek 's pit (amphioxus). It extends from the surface ectoderm to a position ventral to the prosencephalon. The external opening of the pouch is the nostril, on the dorsal midline.
Lampreys are monorhine agnaths which are unusual among vertebrates in possessing of a single olfactory capsule rather than the more typical pair of capsules. This single capsule is located on the posterior wall of the nasohypophyseal pouch. The olfactory capsule is the sense organ associated with the prosencephalon.
Lampreys, like many early vertebrates have three eyes consisting of a pair of lateral eyes on the sides of the head and a single, unpaired median eye on top of the head. Each lateral eyeis a spherical black structure lying beside the posterior prosencephalon. Its position marks the boundary between the prosencephalon and mesencephalon. It is however, functionally associated with the mesencephalon but lies beside and is physically connected to the diencephalon of the prosencephalon.
The inner ear (otic vesicle) is the sense organ associated with the rhombencephalon and it can be seen as a large clear oval beside the anterior rhombencephalon. It lies just posterior to the division between the mesencephalon and rhombencephalon. During embryonic development the olfactory, optic, and otic sense organs develop as invaginations of surface ectoderm.
The notochord is a long thick rod lying ventral to the neural tube. It extends from the level of the inner ear posteriorly to the tip of the tail. Note that the notochord extends anteriorly only as far as the mesencephalon. Compare its length with that of the notochord of amphioxus. The name “cephalochordate” for amphioxus alludes to the presence of the notochord in the entire length of the head.
The posterior wall of the vestibule is a muscular partition, the velum. The gut begins with the mouth which penetrates the velum under the oral hood. The mouth is surrounded by smallbuccal cirri and cannot be seen in lateral view. Posterior to the mouth are visible the muscular lateral folds of the velum. Movements of the velum help move water through the mouth into the pharynx.
The pharynx with its seven visceral (= gill, = branchial) pouches is the region of the gut posterior to the mouth. Separating the pouches are eight visceral arches which bear gill lamellae on their anterior and posterior walls. Ammocoetes, like amphioxus, is a filter feeder, but it uses the muscular velum, rather than cilia, to generate a flow of water that is the respiratory and feeding current. The ciliary current of amphioxus is chiefly for feeding and is not important in respiration. Also unlike amphioxus, the gill pouches open directly to the exterior instead of into an atrium and the gills function both in feeding and in respiration.
The endostyle lies ventral to the anterior portion of the pharynx. It is homologous to the endostyle of the protochordates and part of it will develop into the thyroid gland of the adult lamprey. It has an endocrine function and is homologous to the thyroid of other vertebrates.
During metamorphosis the lumen of the pharynx separates from the lumen of the gut and becomes a blind pouch. In adults it serves a strictly respiratory function, no longer being for filterfeeding by the parasitic adults. Food (the prey’s blood) flows posteriorly through the esophagus whereas a tidal respiratory current enters and leaves the gill pouches via the external gill slits. The mouth is permanently occluded by its attachment to the prey and cannot be used as an intake for the respiratory current.
The narrow esophagus leads posteriorly from the pharynx and soon widens to become the intestine. This portion of the gut extends posteriorly to open to the exterior via the anus. Use the 10X lens to examine the gut contents. You should be able to find the siliceous frustules (cell walls) of diatom (algae) the ammocoetes has filtered from the water.
Examine the area between the pharynx and intestine using the 10X lens. Immediately posterior to the pharynx is the heart lying ventral to the esophagus. The liver lies just behind the heart and is also ventral to the esophagus. A large, easily seen, spherical, clear gall bladder lies near the liver. The gall bladder is lost during metamorphosis and is not present in the adult. In some specimens it may be possible to see the very faint fingerlike tubules of the pronephros by focusing carefully in the region posterior to the heart and above or below the esophagus. The pronephros is the kidney of the larval cyclostome. It will be replaced by a mesonephros in the adult lamprey.
The body musculature is divided into overlapping myomeres. The muscle masses can be seen most clearly as a band running longitudinally just below the dorsal fin and above the nerve cord. Of course, they extend over the lateral aspects of the animal also but cannot be seen in that region because of their transparency. Look carefully in the posterior region of the animal for the myosepta that separate successive myomeres.
Preserved Specimen
Use the dissecting microscope to examine a large preserved specimen. It should be about 100 mm in length and should be immersed in water in a small dissecting pan. Use microdissecting forceps to manipulate the larva as needed. These specimens are fragile but are used year after year. Please handle them gently.
Head
The body consists of a head, trunk, and tail. The head is small and poorly demarcated from the much longer trunk. It, the head, extends posteriorly from the pointed snout to about the level at which the body achieves its maximum diameter. At the anterior end of the head a large oral hood overhangs the spacious cavity known as the vestibule (Fig 1). During development the oral hood expands to form the walls of the adult buccal funnel. The posterior-lateral corners of the hood are free oral lobes, attached only at their bases. The inner roof of the vestibule is covered by patches buccal cirri (= oral papillae), which are easily seen with magnification. The posterior wall of the vestibule is the velum, a vertical, muscular, transverse partition covered by larger buccal cirri. The cirri play a role in selecting or rejecting food items by the filter feeding ammocoetes. They are both mechanical and sensory. The large mouth is an opening in the center of the velum. It is difficult to see because it is covered and obscured by the large buccal cirri. Externally a pair of inconspicuous oblique folds on the side of the head, the velar folds, indicate the location of velar muscles of the velum. The mouth is not supported by jaws in these or any other agnath (= without jaws). The upper lip is the median dorsal portion of the oral hood. The lower lip extends as a low transverse ridge between the oral lobes of the hood.
The nostril is a conspicuous pore on the dorsal midline of the head. It is completely surrounded by a circular lip. Note that there is only one nostril in these monorhine (= one nose) agnaths. The nostril opens into the nasohypophyseal pouch which extends deep into the head and curves posteriorly ventral to anterior brain (prosencephalon). The pouch is not visible in these opaque specimens. The nasohypophyseal pouch is of evolutionary interest as the homolog of the pituitary gland (= hypophysis). On its posterior wall is a single olfactory sac bearing the highly folded olfactory epithelium responsible for distance chemoreception (= olfaction or smell).
Anterior to the nasohypophyseal pore are two short dorso-lateral rows of small lateral line pores. These are openings of the lateral line system, the all important sensory system of fishes. Other lateral line pores can be seen posterior to the nostril and at other places, especially on the head. Two long dorso-lateral rows of widely spaced pores extend along the trunk, near the dorsal midline.
The two lateral eyes are on the sides of the head a little posterior to the level of the nostril but are deeply insunk into the head and are nonfunctional in the ammocoetes. You will not see them because they are deep in the opaque, uncleared tissue of the head. The unpaired third, or pineal, eye is on the dorsal midline immediately posterior to the nostril but is not discernable.
Trunk
Most of the length of the body is the trunk. It extends from the anterior end of the widest part of the body posteriorly to the anus. The long dorsal fin extends along the dorsal midline for the entire length of the trunk (Fig 1). The dorsal fin is continuous with the caudal fin of the tail.
The swollen anterior portion of the trunk is the branchial region (= pharyngeal region). It contains the pharynx, seven pairs of gill pouches, seven pairs of external gill slits, and eight paired gill arches characteristic of lamprey larvae and adults. The seven external gill slits are easily seen forming a short line along both sides of the branchial region. Each gill slit is a more of less circular pore (which is probably closed) posterior to a U-shaped depression in the body wall, which may superficially appear to be another gill slit. It is not, however, rather it is a thin flap of tissue serving as a one-way, no-return valve to prevent the entry of water from the outside into the external gill slit. With its tip pointed anteriorly, gently insert a microteasing needle into an external gill slit, under the valve, and use it to lift the valve slightly. This will open the slit revealing the lumen of the gill pouch from which the slit opens. The internal gill slit and pharynx cannot be seen without dissection, which you should not attempt with these specimens. The gill lamellae are located on the anterior and posterior walls of the pouches but cannot be seen without dissection. The highly vascularized convoluted lamellae are the sites of gas exchange.
Pharyngeal muscles, assisting the velar muscles mentioned above, pump water into the mouth and pharynx (Fig 1). In amphioxus the feeding current is generated by cilia but in the ammocoetes muscles are used to generate a current that is both feeding and respiratory. Food particles in the incoming water remain in the pharynx and pass posteriorly into the intestine for digestion and absorption whereas the water passes laterally into an internal gill slit, then into a gill pouch where gas exchange occurs, and then out the external gill slit. Note that the ammocoetes larva has no atrium and no atriopore and in this regard differs from amphioxus. The ciliary currents suitable for small animals are unable to create a water current sufficient to supply larger animals with food and oxygen. Substitution of muscular for ciliary pumps is one of the innovations that made it possible for vertebrates to become large.
The remainder of the trunk is smaller in diameter than the branchial region. The vent on the midventral line near the termination of the caudal fin marks the posterior limit of the trunk. The vent is the external opening of the cloacal pit. The intestine and kidney release their wastes into the cloaca. The vent is flanked by a pair of lips. Insert the microteasing needle between the lips to reveal the shallow cloacal pit. The anterior opening into the pit is the anus of the digestive system and the posterior opening is the urinary pore of the excretory system. The urinary pore sits atop a small urogenital papilla.
Observe the sides of the trunk (and tail) noting the segmentally arranged bands of axial myomeres (musculature) separated by connective tissue myosepta. The ventral midline of the trunk is marked by a longitudinal ridge, which shows no signs of segmentation. This is the transverse muscle (Fig 2). Metapleural folds are absent.
Note the absence of paired appendages, there being neither pectoral nor pelvic fins in the agnath fishes. Jaws are also absent but that is not obvious externally. The absence of jaws and paired appendages are agnath apomorphies.
Tail
The tail is defined as the region of the body posterior to the anus (Fig 1). It bears the caudal fin on its midline, dorsal and ventral. The caudal fin is continuous dorsally with the dorsal fin but ventrally it ends a little posterior to the anus and the ventral trunk has no fin. The postanal tail is a chordate apomorphy.
Return the specimen to the jar of preservative when finished.
Pharyngeal Cross Section
Carefully examine the sections on your slide and select the one through the pharyngeal region for further study. The appearance of your pharyngeal section will vary greatly depending on the level of the visceral pouch sectioned. It may or may not closely resemble Figure 2.
Note first the heavy integument (= skin) surrounding the animal. It is composed of a thick outer epidermis underlain by a thinner connective tissue dermis. Dorsally there is a large mass of axial muscle subdivided into myomeres. These subdivisions overlap each other and are separated by thin connective tissue myosepta. The large vacuolated notochord is located between the right and left muscle masses and is surrounded by a cartilaginous notochordal sheath. Above it is the flat nerve cord (or rhombencephalon depending on the level of the section). The nerve cord is loosely enclosed by a neural sheath of fibrous connective tissue. Anterior sections may include the cartilagenous neurocranium, or braincase, ventral to and lateral to the notochord.
Most of the section is occupied by the enormous pharynx, whose wall is composed of visceral arches. The visceral arch (= branchial arch, gill arch) is an important feature of vertebrate anatomy. The arches are features of the pharynx and they, ot their derivatives are present in all vertebrates. A typical arch a U-shaped structure consisting of a skeleton, a cranial nerve, an aortic arch, muscles, and gill lamellae. Collectively the series of arches form the wall of the pharynx and the open spaces between successive arches are the gill pouches.
The open space in the center of the section is the lumen of the pharynx. It is flanked by a visceral arch (or arches). The number visible depends on the level of section and the degree of distortion of these preserved specimens. If the plane of section passes between two successive arches then the arch will not be seen, at least not as a continuous U. The right and left limbs of the arch each bear numerous horizontal gill filaments, which may be attached by either their lateral or medial ends. The filaments protrude into the visceral pouch. They bear tiny fingerlike papillae, the gill lamellae, on their margins. The lamellae are heavily vascularized by branches of an aortic arch and their function is to increase the surface area available for gas exchange. .
Figure 2. Ammocoetes larva. Cross section through the pharynx at 50X. fish160L.gif
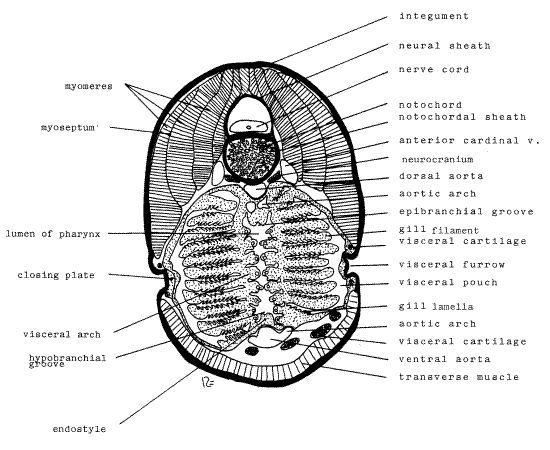
A ciliated longitudinal furrow, the epibranchial groove (epi= above, branchial = gill) is located dorsal to the pharyngeal lumen on the middorsal line while a similar hypobranchial groove (hypo = below) is found ventral to the lumen on the midventral line. The hypobranchial groove lies on the upper surface of the endostyle.
Laterally the body wall of the branchial region is invaginated to form shallow visceral furrows which communicate with the visceral pouch by small circular external gill slits. The slits themselves are small and most sections will not pass through them. In most cases you will see the closing plate of ectoderm and endoderm separating the visceral furrow from the visceral pouch.
Sections through several of the cartilages of the visceral skeleton can be seen ventral and lateral to the pharynx. A thick transverse muscle extends across the ventral aspect of the animal.
A large anterior cardinal vein is located on each side of the notochord. These are important veins draining venous blood from the head to the heart. Unlike amphioxus, the ammocoetes larva has a heart. The single median dorsal aorta is located ventral to the notochord. This large vessel drains oxygenated arterial blood from the gills and delivers it to the head and body.
Ventral to the pharynx is the large ventral aorta. Blood exits the heart via the ventral aorta which runs the length of the pharynx. Paired aortic arches exit the ventral aorta and extent through the gill pouches to the dorsal aorta. En route they break up into capillary beds in the gills where their blood is oxygenated. In some sections the connection between the ventral aorta (Fig 2) or the dorsal aorta may be visible.
Other
Examine the demonstration of the brook lamprey life cycle is one is available.
References
Feduccia A . 1975. Structure and Evolution of Vertebrates. Norton. New York. 168p.
Hyman LH . 1942. Comparative Vertebrate Anatomy. Univ. Chicago
Press. Chicago. 544 p.
Lehman HE . 1983. Chordate Development. Hunter. Winston-Salem, North
Carolina. 342p.
Ruppert EE, Fox RS, Barnes RB. 2004. Invertebrate Zoology, A functional evolutionary approach, 7 th ed. Brooks Cole Thomson, Belmont CA. 963 pp.
Weichert CK . 1961. Representative Chordates. McGraw-Hill. New York.
216 p.
Supplies
Compound microscope
Ammocoetes larva wholemount
Ammocoetes larva cross section slide through pharynx
Dissecting microscope
Large preserved ammocoetes, about 100 mm
Small dissecting pan (an empty kippered herring snack tin is ideal)
Microteasing needle and microdissecting forceps.